La sérotonine est un produit chimique qui transporte des messages entre les cellules nerveuses du cerveau et dans tout votre corps. La sérotonine joue un rôle clé dans des fonctions corporelles telles que l’humeur, le sommeil, la digestion, les nausées, la cicatrisation des plaies, la santé des os, la coagulation sanguine et le désir sexuel. Des niveaux de sérotonine trop faibles ou trop élevés peuvent entraîner des problèmes de santé physique et psychologique.
Qu’est-ce que la sérotonine ?
La sérotonine, également connue sous le nom de 5-hydroxytryptamine (5-HT), est un neurotransmetteur monoamine. Il agit également comme une hormone.
En tant que neurotransmetteur, la sérotonine transporte des messages entre les cellules nerveuses de votre cerveau (votre système nerveux central) et dans tout votre corps (votre système nerveux périphérique). Ces messages chimiques indiquent à votre corps comment fonctionner.
La sérotonine joue plusieurs rôles dans votre corps, notamment en influençant l’apprentissage, la mémoire, le bonheur ainsi qu’en régulant la température corporelle, le sommeil, le comportement sexuel et la faim. On pense que le manque de sérotonine joue un rôle dans la dépression , l’anxiété , la manie et d’autres problèmes de santé.
La majeure partie de la sérotonine présente dans votre corps se trouve dans votre intestin (intestins). Environ 90 % de la sérotonine se trouve dans les cellules tapissant votre tractus gastro-intestinal. Il est libéré dans votre circulation sanguine et absorbé par les plaquettes. Seulement environ 10 % sont produits dans votre cerveau.
La sérotonine est fabriquée à partir du tryptophane, un acide aminé essentiel. Un acide aminé essentiel signifie qu’il ne peut pas être fabriqué par votre corps. Il doit être obtenu à partir des aliments que vous consommez.
Que fait la sérotonine dans mon corps ?
La sérotonine joue un rôle dans de nombreuses fonctions de votre corps :
Humeur : La sérotonine dans votre cerveau régule votre humeur. On l’appelle souvent le produit chimique naturel du « bien-être » de votre corps. Lorsque la sérotonine est à des niveaux normaux, vous vous sentez plus concentré, émotionnellement stable, plus heureux et plus calme. De faibles niveaux de sérotonine sont associés à la dépression. De nombreux médicaments utilisés pour traiter l’anxiété, la dépression et d’autres troubles de l’humeur visent souvent à augmenter le niveau de sérotonine dans votre cerveau.
Digestion : La majeure partie de la sérotonine de votre corps se trouve dans votre tractus gastro-intestinal, où elle aide à contrôler votre fonction intestinale et joue un rôle dans la protection de votre intestin. Votre intestin peut augmenter la libération de sérotonine pour accélérer la digestion et débarrasser votre corps des aliments irritants ou des produits toxiques. La sérotonine joue également un rôle dans la réduction de votre appétit en mangeant.
Nausée : la nausée se déclenche lorsque la sérotonine est libérée dans votre intestin plus rapidement qu’elle ne peut être digérée. Le message chimique est reçu par votre cerveau, que vous percevez comme une nausée. De nombreux médicaments utilisés pour réduire les sensations de nausée et de vomissement ciblent des récepteurs de sérotonine spécifiques dans votre cerveau.
Sommeil : la sérotonine, associée à un autre neurotransmetteur dopamine, joue un rôle dans la qualité de votre sommeil (la qualité et la durée de votre sommeil). Votre cerveau a également besoin de sérotonine pour fabriquer de la mélatonine, une hormone qui régule votre cycle veille-sommeil.
Cicatrisation des plaies : la sérotonine est libérée par les plaquettes présentes dans votre sang pour aider à cicatriser les plaies. Cela provoque également le rétrécissement des plus petits vaisseaux sanguins, les artérioles, ce qui ralentit le flux sanguin et favorise la formation de caillots. Il s’agit d’un processus important dans la cicatrisation des plaies.
Santé des os : les niveaux de sérotonine peuvent jouer un rôle dans la densité de vos os. Des niveaux élevés de sérotonine dans votre intestin peuvent jouer un rôle dans la faiblesse des os, ce qui peut entraîner des fractures osseuses et l’ostéoporose .
Santé sexuelle : la sérotonine joue également un rôle, avec le neurotransmetteur dopamine, dans votre désir sexuel.
Quels problèmes sont associés à de faibles niveaux de sérotonine ?
De faibles niveaux de sérotonine peuvent être associés à de nombreux problèmes de santé, notamment :
Les scientifiques ont encore beaucoup à apprendre sur le rôle de la sérotonine dans l’organisme et dans les maladies.
Qu’est-ce qui peut causer de faibles niveaux de sérotonine ?
Un faible taux de sérotonine a généralement plusieurs causes. Techniquement, les niveaux de sérotonine sont faibles parce que :
Votre corps ne produit pas suffisamment de sérotonine.
Votre corps n’utilise pas efficacement la sérotonine. Cela peut se produire si vous n’avez pas suffisamment de récepteurs de sérotonine ou si ceux-ci ne fonctionnent pas comme ils le devraient.
Que puis-je faire pour augmenter les niveaux de sérotonine ?
Les moyens d’augmenter les niveaux de sérotonine comprennent :
Manger plus d’aliments contenant du tryptophane.
Obtenir plus de soleil.
Prendre certains suppléments.
Faire plus d’exercice et réduire votre niveau de stress.
Aliments pour augmenter les niveaux de sérotonine
De nombreux aliments contiennent naturellement du tryptophane, l’acide aminé à partir duquel la sérotonine est fabriquée. Vous pouvez essayer d’augmenter votre taux de sérotonine en mangeant des aliments contenant du tryptophane, tels que :
Saumon.
Œufs.
Fromage.
Dinde.
Tofu.
Ananas.
Noix, avoine et graines.
Manger des aliments riches en tryptophane n’augmentera pas nécessairement les niveaux de sérotonine. C’est un processus complexe. Votre corps a besoin de glucides pour libérer l’insuline, nécessaire à l’absorption des acides aminés. Même si le tryptophane pénètre dans votre sang, il doit entrer en compétition avec d’autres acides aminés pour être absorbé par votre cerveau. Les scientifiques étudient encore comment la consommation d’aliments contenant du tryptophane pourrait augmenter les niveaux de sérotonine.
Lumière du soleil
Ne pas être suffisamment exposé au soleil peut entraîner des troubles de l’humeur et des troubles affectifs saisonniers chez certaines personnes. Essayez de profiter de 10 à 15 minutes de soleil chaque jour pour augmenter non seulement les niveaux de sérotonine, mais également les niveaux de vitamine D. Si vous vivez dans une région où vous ne pouvez pas obtenir la lumière naturelle du soleil, envisagez d’utiliser la luminothérapie pour obtenir la lumière du soleil quotidienne dont vous avez besoin.
Suppléments
Plusieurs compléments alimentaires et à base de plantes augmentent également les niveaux de sérotonine. Ceux-ci inclus:
Complément alimentaire : Tryptophane, probiotiques et SAMe.
Suppléments à base de plantes : ceux-ci peuvent inclure le ginseng, le millepertuis , la rue syrienne et la muscade.
Exercice
On sait que l’exercice régulier augmente les niveaux de sérotonine. Trente minutes d’ exercices aérobiques cinq fois par semaine et deux séances de musculation par semaine peuvent améliorer les troubles de l’humeur et la santé cardiaque.
Quels médicaments augmentent les niveaux de sérotonine ?
La sérotonine ou les récepteurs de la sérotonine sont des cibles courantes de l’industrie pharmaceutique puisque de nombreux problèmes de santé sont affectés par la sérotonine. Certains des médicaments les plus courants qui augmentent les niveaux de sérotonine sont les suivants.
Plusieurs classes différentes d’ antidépresseurs bloquent la réabsorption et le recyclage de la sérotonine, permettant ainsi à une plus grande quantité d’en rester dans le cerveau. Les médicaments qui agissent de cette façon sont les inhibiteurs sélectifs de la recapture de la sérotonine (par exemple, la paroxétine [Paxil®]), les inhibiteurs de la recapture de la sérotonine et de la noradrénaline (par exemple, la venlafaxine [Effexor®]) et les antidépresseurs tricycliques (par exemple, l’amitriptyline [Elavil®]). Un autre type d’antidépresseur, les inhibiteurs de la monoamine oxydase (par exemple, la phénelzine [Nardil®]) bloque une enzyme qui décompose la sérotonine.
De nombreux autres médicaments, pris pour de nombreuses pathologies, augmentent également la sérotonine. Certains de ces médicaments comprennent la famille des triptans, des analgésiques opioïdes, des antitussifs contenant du dextrométhorphane et des médicaments contre la nausée.
Quels problèmes sont associés à des niveaux élevés de sérotonine ?
Le syndrome sérotoninergique est une maladie qui survient lorsque les niveaux de sérotonine augmentent trop. Cela se produit généralement si vous augmentez la dose d’un médicament connu pour augmenter les niveaux de sérotonine ou si vous prenez un autre médicament connu pour augmenter la sérotonine.
Les symptômes légers comprennent des frissons, une transpiration abondante, de la confusion, de l’agitation, de l’hypertension artérielle, des contractions musculaires et de la diarrhée. Les symptômes graves comprennent une forte fièvre, des convulsions, des évanouissements et un rythme cardiaque anormal.
Le syndrome sérotoninergique peut être mortel s’il est grave et s’il n’est pas détecté tôt et traité rapidement.
Questions courantes supplémentaires
Quelle est la différence entre la dopamine et la sérotonine ?
La dopamine et la sérotonine sont toutes deux des neurotransmetteurs. Cela signifie qu’ils véhiculent des messages chimiques entre les cellules nerveuses du cerveau ainsi que vers et depuis d’autres zones de votre corps. Les deux sont également considérées comme les « hormones du bonheur », car elles jouent toutes deux un rôle dans l’humeur et les émotions positives. La sérotonine est associée au bonheur, à la concentration et au calme. La dopamine est associée aux récompenses et à la motivation. La dopamine et la sérotonine sont également impliquées dans certains problèmes de santé mentale, notamment la dépression et les troubles de l’humeur.
La dopamine et la sérotonine ont également des fonctions distinctes. La dopamine contrôle les mouvements et la coordination du corps. La sérotonine aide à réguler les fonctions digestives, notamment la fonction intestinale et l’appétit. La dopamine provoque une sensation de faim tandis que la sérotonine supprime cette sensation. La dopamine est principalement stockée dans votre cerveau tandis que la sérotonine se trouve principalement dans votre intestin.
Parfois, ces neurotransmetteurs travaillent ensemble pour maintenir un équilibre chimique minutieux dans votre corps. Parfois, un déséquilibre entraîne une surproduction de l’autre neurotransmetteur. Avoir trop ou pas assez de l’un ou l’autre peut provoquer des symptômes physiques et psychologiques.
Un message de la Cleveland Clinic
La sérotonine joue un rôle clé dans des fonctions corporelles telles que l’humeur, le sommeil, l’appétit, l’anxiété, la digestion, la coagulation sanguine et le désir sexuel. Si vous souffrez d’un problème de santé qui affecte la sérotonine ou qui est affecté par la sérotonine, demandez à votre médecin ce que vous devez savoir sur la sérotonine. Pour éviter une variation du taux de sérotonine de votre corps et réduire le risque d’interactions médicamenteuses, n’arrêtez pas de prendre vos médicaments, ne modifiez pas votre dose et ne prenez pas de suppléments alimentaires ou à base de plantes sans en parler au préalable avec votre fournisseur.
On a encore beaucoup à découvrir dans de si nombreux domaines comme la santé humaine , mais aussi la santé de la vie du sol et la santé des plantes ……A quel niveau intervient l’action des rayons solaires sur cette santé commune à tous, sachant que le soleil nous fait bénéficier de son rayonnement depuis un bon moment …..Sans soleil , sous une serre agricole , la qualité de l’alimentation est-elle la même …..??
Préface : Lorsque j’en ai écrit la version originale en 2006, je l’ai montrée à un ami. Il m’a demandé si j’avais lu les Adieux à la nation de George Washington. Je n’avais pas. En lisant son avertissement aux Américains en 1796, j’ai réalisé qu’il avait précédé ma réflexion de plusieurs siècles et je lui attribue donc l’un des points majeurs de cet essai. Bien que je n’écrive pas sur l’Amérique, mais plutôt sur la nation de ma naissance – le Zimbabwe – l’avertissement de Washington sur les dangers d’autoriser les partis politiques est évident pour toutes les personnes bonnes et intelligentes d’aujourd’hui en Amérique. Comme c’est mon habitude de toujours, je m’attarde non pas sur les problèmes, mais sur les solutions.
Ces réponses à la première expression de mes pensées par des personnes réfléchies m’ont incité à la mettre à jour et à la publier sur mon blog non censuré, comme je le fais actuellement.
Réactions à la bonne gouvernance.
Toutes nos félicitations! La « Bonne gouvernance en Afrique » est un chef-d’œuvre, une pépite d’or pur. Je parie qu’avec le temps, il prendra rang parmi les grands ouvrages du genre du gouvernement et de la philosophie. Vous avez atteint le point mort cible. Sans grandiloquence ni pompe, vous avez traité un sujet délicat de manière astucieuse et érudite. C’est un article bien écrit ; clairement compréhensible et pertinent, qui transmet également un message puissant. Sam Brown, Colorado.
OUAH…..!!! Merci beaucoup pour ce document remarquable et réfléchi. C’est véritablement révolutionnaire – dans le meilleur sens du terme. Bien entendu, ce ne sont pas seulement les Zimbabwéens qui ont besoin de lire ceci, et il devrait être publié quelque part où il sera largement lu et stimulera un véritable débat à grande échelle. Je ne crois pas avoir jamais vu une explication aussi convaincante de la raison pour laquelle les gouvernements du monde entier sont incapables d’élaborer une politique environnementale sensée, et votre démolition systématique du pouvoir des partis est si importante à une époque où la démocratie et la politique des partis sont considérées comme indissociables. Avez-vous des projets de publication ? Je vous inviterais à rédiger une version courte – peut-être plus générique (c’est-à-dire avec moins de références zimbabwéennes détaillées) – et à la soumettre à quelque chose comme le New York Times Magazine, par exemple.
Ron Coleman, Canada.
En effet, il s’agit d’une référence en matière d’essais politiques et de gestion !! Puis-je le partager avec quelques collègues de l’École de communication ? L’un est mon penseur « radical » et politique local préféré, et l’autre est un extraordinaire catalyseur de l’éducation transdisciplinaire.
Dick Richardson, Université du Texas, Austin.
Bonne gouvernance en Afrique.
Par Allan Savory.
Introduction
Le Zimbabwe est en difficulté après avoir répété le modèle d’autres États africains après l’indépendance. Mais considérez ceci : les grands châteaux de Grande-Bretagne n’ont été construits qu’après la fin de la colonisation romaine, lorsque les seigneurs de guerre anglais se battaient pour la suprématie et que les rois assassinaient frères, épouses et autres personnes au fil des siècles pour atteindre le même objectif. Ce n’est que lentement et douloureusement que le peuple britannique a réussi à combler le fossé entre les dirigeants tyranniques et les dirigeants démocrates pour jouir des libertés démocratiques dont lui et les millions d’immigrants des anciennes colonies jouissent aujourd’hui. En Afrique, nous essayons de combler cet écart en quelques décennies, et à une époque où les défauts des démocraties occidentales conduisent de plus en plus à une corruption flagrante et à une dégradation de l’environnement que peu de gens associeraient à leurs systèmes politiques. Et pourtant, ils sont étroitement liés.
Tant que tous les gens ne se sentiront pas libres, en sécurité et bien gouvernés, aucun ne le sera. Des terres pauvres conduisent inévitablement à la pauvreté, à l’effondrement social, à la pauvreté, à la violence, à l’instabilité politique et, trop souvent, au génocide. Ces deux convictions ont dominé ma vie d’adulte en tant que scientifique africain indépendant de quatrième génération né au Zimbabwe. De telles convictions m’ont conduit brièvement à la vie politique, puis à l’exil. Alors que le lien entre la santé de la terre et la stabilité politique, sociale et économique a été nié pendant des années par la plupart des pays, il est désormais de plus en plus reconnu. Pour l’Afrique, il est important de reconnaître que la santé, la stabilité et la productivité de nos terres sont aussi fondamentales pour un gouvernement stable que la justice sociale.
En tant qu’ancien allié politique de Mugabe, Nkomo, Tongogara, Zvobo, Edson Sithole, Dabengwa, Chinamano et de nombreux autres Zimbabwéens de toutes couleurs et tribus dans notre lutte pour la démocratie et l’indépendance, je sais que nos rêves se sont égarés. Je sais aussi que lorsque nous avons travaillé ensemble en Suisse vers les dernières étapes de notre longue guerre, nous étions simplement des Zimbabwéens, sans distinction de couleur ou de tribu, avec un objectif commun : obtenir notre indépendance en tant que nation fière et démocratique. Nous avons fait heurter notre navire contre les rochers pour de nombreuses raisons, dont certaines sont indépendantes de notre volonté, comme je vais l’expliquer. Aujourd’hui, une fois de plus, nous avons un objectif commun : travailler ensemble pour faire décoller notre navire d’État des rochers, le redresser et flotter fièrement à flot. Nous ne sommes pas une nation de mendiants et nous pouvons assurer un leadership pour l’Afrique et au-delà.
Bien que riches en ressources, de nombreux pays africains sont si délabrés financièrement et dépendants de l’aide étrangère qu’ils sont difficilement indépendants. Même si les dirigeants et les partis politiques qui ont conduit leurs pays à l’indépendance ont généralement été blâmés, je pense que ce blâme est déplacé. Alors qu’un sort similaire est arrivé à la plupart des jeunes démocraties africaines et que le changement de parti au pouvoir a, au mieux, abouti à une amélioration marginale de la vie des gens avec une dépendance continue à l’égard de l’aide étrangère, cela suggère qu’il pourrait y avoir une cause plus profonde aux mauvais résultats de la démocratie en Afrique. Afrique. Le fait que tant de pays aient connu les mêmes problèmes et que même la puissance économique de l’Afrique du Sud s’engage sur la voie du Zimbabwe suggère qu’il pourrait y avoir quelque chose qui ne va pas dans le système démocratique hérité des nations africaines. Blâmer les individus ou les partis au pouvoir, plutôt que d’examiner les causes profondes, n’est ni constructif ni susceptible de conduire à une bonne gouvernance.
Les opinions que j’exprime sont en gestation depuis plus de cinquante ans et en particulier dans les presque quarante années qui ont suivi mon mandat au Parlement. Je suis apolitique et je suis entré au Parlement en désespoir de cause pour lutter contre le racisme, les politiques destructrices de l’environnement et pour tenter de mettre fin à une guerre insensée. Au parti d’opposition que j’ai ensuite dirigé, j’ai toujours souligné que je n’étais qu’un leader en temps de guerre et que je me retirerais de la politique dès que nous pourrions mettre fin à la guerre et obtenir notre indépendance. La raison de mon refus, malgré les demandes, de continuer en politique était simplement que je savais qu’assurer une bonne gouvernance dépassait mes capacités ou ma compréhension. Il m’a fallu quarante ans pour comprendre ce qui empêche la bonne gouvernance dans un pays et donc ce qui pourrait être fait pour y parvenir.
Dans cet article, je présente de nouvelles idées managériales et scientifiques qui sont contre-intuitives ou qui changent de paradigme, et qui expliquent pourquoi il s’est avéré si difficile pour un gouvernement, quelle qu’en soit la forme, d’assurer une bonne gouvernance. Et j’explique les lacunes du système de conviction politique hérité du Zimbabwe ainsi que les structures parlementaires et de la fonction publique qui en découlent. Ces lacunes ont rendu inévitables les troubles rencontrés dans les jeunes démocraties africaines – quel que soit celui qui les dirigeait. Le Zimbabwe ne peut pas se sortir de ses difficultés, quelles que soient les bonnes intentions de ses dirigeants politiques actuels ou futurs, à moins que les Zimbabwéens ne réfléchissent à nouveau.
Je crois sincèrement que les suggestions que je propose ont le potentiel, dans le cas du Zimbabwe, de produire rapidement une gouvernance supérieure à celle des démocraties plus anciennes, et dont elles pourraient tirer des leçons. Ces suggestions pourraient conduire à une solution interne au Zimbabwe dans laquelle il y aurait pour la plupart des gagnants et peu de perdants et qui pourrait être adoptée par la plupart de mes compatriotes. Tout en me concentrant sur mon propre pays, je suis conscient que d’autres pays, comme l’Afrique du Sud, la Zambie, la Namibie et bien d’autres encore, pourraient adopter tout ce que j’écris pour leur permettre également de jouir d’une bonne gouvernance dans laquelle tous se sentent en sécurité et bien gouvernés.
Les idées que j’exprime ne visent pas à offenser les individus d’un parti politique, mais sont proposées dans l’espoir d’encourager une discussion ouverte et nouvelle pour nous aider à nous diriger vers un avenir meilleur et à le faire rapidement.
Tout en m’intéressant à l’Afrique, et en particulier au Zimbabwe, j’établis des parallèles avec les États-Unis et d’autres pays quant aux leçons que nous pouvons en tirer. Qu’est-ce qui empêche même les hommes politiques les mieux intentionnés d’assurer une bonne gouvernance constante dans n’importe quelle nation et pas seulement dans la mienne ?
Le niveau de destruction de l’environnement mis en évidence par la désertification mondiale et maintenant le changement climatique mondial, combiné à l’augmentation de la population et des aspirations, exigera un besoin de bonne gouvernance plus grand que jamais dans l’histoire. J’espère que les idées avancées ici encourageront le débat et une nouvelle réflexion dans des pays autres que le Zimbabwe, y compris nos voisins proches, la Zambie et l’Afrique du Sud, et parmi des personnes autres que les politiciens. Tout comme les meilleurs fabricants de bougies n’auraient jamais pu imaginer ou développer l’éclairage électrique, il est également peu probable que les hommes politiques voient clairement les solutions que les gens ordinaires voient avec clarté.
Section I. Principales connaissances managériales et scientifiques
Si, en tant que fondateur et dirigeant d’un parti politique, je n’avais pas été obligé d’élaborer des politiques en articulant le programme d’un parti, je n’aurais jamais compris que le rôle le plus important du gouvernement est la formulation de politiques dont découlent des lois et des réglementations. L’opposition à tout parti politique est invariablement due au fait que les politiques qu’il développe ou qu’il adopte sont considérées comme préjudiciables à ceux qui proposent d’autres politiques encore.
Si ces politiques ont un impact sur tous les domaines de notre vie – citoyenneté, fiscalité, éducation, etc. – ce sont les politiques environnementales qui nous impactent le plus profondément à long terme.
Les politiques environnementales affectent directement l’économie et la qualité de vie des gens, ce qui à leur tour détermine s’ils vivent en paix ou, à terme, dans le chaos et le génocide.
Avant de discuter d’une meilleure forme de gouvernance pour le Zimbabwe, nous devons examiner une nouvelle compréhension des cinquante dernières années qui explique pourquoi la bonne gouvernance échappe à toutes les formes de gouvernement dans n’importe quelle région du monde. De nombreux facteurs sont faussement imputés aux échecs des gouvernements et de leurs politiques, mais nous devons y regarder plus en profondeur : nous devons comprendre le rôle de la gestion, qui inclut les projets de développement et l’élaboration de politiques.
Bien qu’il existe de nombreuses écoles de gestion, consultants et experts conseillant les gouvernements, à ma connaissance, aucun n’a jamais étudié le cadre fondamental de gestion génétiquement intégré chez tous les animaux utilisant des outils, y compris les humains. Un cadre reconnaissable du ménage à la gouvernance car c’est universellement la manière dont les humains ont toujours pris des décisions en matière de gestion pour améliorer nos vies. Pour comprendre rapidement ce que je dis, pensez à ce que la société, et donc les scientifiques et les gouvernements, blâment pour la désertification mondiale et le changement climatique. Nous blâmons plus que toute autre chose, l’élevage, le charbon et le pétrole.
Cependant, le bon sens nous dit qu’il s’agit de ressources et qu’aucune ressource ne peut poser de problème. La seule chose qui peut provoquer la désertification mondiale et le changement climatique est la gestion de ces ressources. C’est la gestion au fil des siècles qui a conduit à ce que l’élevage soit à l’origine des grands déserts de l’Antiquité. C’est la direction qui choisit d’utiliser le charbon et le pétrole comme combustibles fossiles et de les brûler à un rythme préjudiciable à l’environnement. C’est donc incontestablement une gestion à 100 % qui est à l’origine des maux de la société – la gestion à tous les niveaux, y compris l’élaboration des politiques.
Si nous ne sommes pas conscients que la gestion est à l’origine des problèmes, ni conscients de ce qui est défectueux dans la gestion, c’est, je crois, dû à l’effet masquant du succès technologique. Si nous regardons tout ce que nous fabriquons ou construisons – des brosses à dents aux ordinateurs, en passant par les satellites, les églises et les hôpitaux, l’électricité à partir du charbon ou du vent et du soleil, etc., tout ce que nous fabriquons, que voyons-nous ? Nous ne pouvons rien faire sans expertise et sans technologie sous une forme ou une autre. Je n’ai aucune idée de comment fabriquer ne serait-ce qu’une simple montre ou un pardessus. Rien de ce que nous fabriquons n’est auto-organisé et capable de continuer lorsqu’une pièce est cassée, manquante ou qu’une source d’énergie (carburant ou batterie) s’épuise. Rien de ce que nous fabriquons, même les véhicules spatiaux, n’implique le concept de complexité, même s’ils peuvent être très compliqués. Avec les choses que nous fabriquons, nous connaissons des succès toujours croissants – il n’y a pas de fin en vue aux merveilles de la technologie et de l’ingéniosité. La plupart des espoirs pour l’avenir reposent sur la technologie et les choses que nous fabriquons.
Examinons maintenant tout ce que nous gérons – l’agriculture, les églises, les économies, les organisations humaines, la gouvernance – la gestion à tous les niveaux, depuis les ménages jusqu’à la gouvernance nationale et les organisations internationales – nous voyons une image différente. Ce que nous gérons n’implique pas toujours la technologie, les organisations et tout ce que nous gérons dans la nature ne cesse pas de fonctionner si une personne meurt ou si une espèce est anéantie. Les choses continuent mais sous une forme modifiée en raison de la complexité auto-organisée. En science des systèmes, tout ce que nous gérons est défini comme un système complexe auto-organisé. C’est dans ce domaine de notre vie – la gestion impliquant une complexité auto-organisée – que nous sommes confrontés à de plus en plus de problèmes, comme un tsunami après l’autre qui nous frappe. Nos succès sont au mieux à court terme et sont trop souvent suivis de conséquences imprévues et de problèmes supplémentaires.
La gestion, et en particulier les projets de développement et l’élaboration de politiques, implique toujours un réseau de complexité sociale/culturelle, économique et environnementale – cette complexité de gestion, comme je l’appellerai, est inévitable depuis la gestion de nos vies jusqu’à la gouvernance. C’est cette complexité des systèmes auto-organisés que nous n’avons jamais été capables de bien gérer.
Il y a une raison simple pour laquelle nous ne gérons pas bien le réseau de la complexité : c’est parce que la gestion est réductionniste – et que le monde réel fonctionne de manière holistique et est complexe. C’est ce que nous avons découvert récemment – au début des années 1980 au Zimbabwe, presque par hasard, comme cela arrive souvent.
J’expliquerai plus tard la différence entre notre gestion universelle et la gestion de la complexité. Mais avant de poursuivre, je dois souligner une autre raison pour laquelle la bonne gouvernance échappe aux gouvernements africains (américains et autres) et elle remonte à l’influence des prix Nobel.
Lorsque l’idée de prestigieux prix Nobel a été évoquée pour la première fois, les meilleurs cerveaux déterminaient quels domaines de l’activité humaine étaient importants pour l’avenir de l’humanité. Décider quels domaines étaient les plus importants pour l’avenir de l’humanité était une tâche extrêmement sérieuse, sachant qu’un tel prestige attirerait les meilleurs esprits vers les domaines les plus importants. Alors que des esprits brillants débattaient sur les mathématiques, aucun ne considérait même l’agriculture ou l’environnement comme importants pour notre avenir. Nous ne voyons donc pas de prix Nobel ni pour l’agriculture ni pour l’environnement, deux domaines plus importants que tous les autres réunis.
L’agriculture n’est pas simplement une production végétale. L’agriculture est la production d’aliments et de fibres à partir des terres et des eaux de la planète. Les cultures, l’élevage, la foresterie, la pêche et la faune sont tous concernés. La quasi-totalité de la planète est impliquée dans l’agriculture, les terres arables constituant environ 20 % de la superficie terrestre, soit 6 % de la surface de la planète – 94 % étant le reste de l’agriculture.
Sans agriculture, nous ne pouvons pas avoir de ville, d’église, d’université, de gouvernement – c’est le fondement de la civilisation, qui est par définition basée sur la ville. Le sort des civilisations suit le sort de l’agriculture.
Le Zimbabwe et les pays voisins ne disposent que d’environ 10 % de terres arables, mais toutes ces terres doivent subvenir aux besoins de la population grâce à l’agriculture au sens le plus large du terme. Pour nous, la seule base durable à long terme de toute économie est le processus photosynthétique – des plantes vertes poussant sur un sol en régénération. Ni le charbon, ni le cuivre, ni l’or, ni les diamants, ni le chrome ne nourrissent les populations, ne préviennent les inondations et les sécheresses, ne luttent contre la désertification ou le changement climatique.
De nombreuses politiques conduisent à divers types de projets de développement. Les politiques et les projets abordent un problème d’une manière ou d’une autre et doivent tous deux être solides.
IA. Les politiques et projets gouvernementaux ne parviennent pas à répondre simultanément aux préoccupations sociales, environnementales et économiques.
Les gouvernements élaborent des politiques pour l’une des deux raisons suivantes : soit pour résoudre un problème, soit pour prévenir un problème prévisible. Pour atteindre son objectif, toute politique (ou projet) doit non seulement s’attaquer à la cause du problème, mais également s’attaquer simultanément à l’ensemble de la complexité sociale/culturelle, environnementale et économique.
L’augmentation massive de la population et la dégradation des terres sont à l’origine de l’essentiel de la pauvreté et de la violence croissante que connaît l’Afrique. La dégradation des terres (désertification) entraîne inévitablement une augmentation de la fréquence et de la gravité des inondations et des sécheresses, sans changement des conditions météorologiques. Et la désertification conduit à la pauvreté, à l’effondrement social, à la violence, à l’instabilité politique, au génocide et à une émigration massive vers les villes et au-delà des frontières (ce qui change même désormais le visage politique de l’Europe). En fait, la désertification est à l’origine de la plupart des symptômes auxquels les gouvernements africains (et maintenant les gouvernements européens) et les agences de développement qui les assistent luttent et dont souffrent et meurent des millions de personnes. On peut s’attendre à ce que tous ces problèmes liés à la désertification s’aggravent au Zimbabwe et dans la majeure partie de l’Afrique, car les derniers objectifs de développement durable (ODD) des Nations Unies s’attaquent presque tous aux symptômes de la désertification et aucun ne s’attaque en aucune façon à la cause.
IB. Apprendre les causes de la désertification a permis de comprendre pourquoi les politiques gouvernementales ne peuvent pas gérer la complexité comme il se doit.
Le travail de ma vie en tant que scientifique a été consacré à percer le mystère de la désertification qui m’a vraiment alarmé pour la première fois alors que j’étais au Sénégal en 1960, représentant le gouvernement rhodésien de l’époque. Ce processus de dégradation des terres, commencé il y a des milliers d’années, a défié nos efforts pour l’inverser et a détruit de nombreuses sociétés et civilisations. Ce que j’ai découvert, c’est que, contrairement à l’opinion dominante, la désertification n’est pas causée par les nombreux facteurs souvent pointés du doigt, tels que le surpâturage, la surpopulation, le surpeuplement du bétail, la propriété communale des terres, etc. Le fait que des États américains entiers, peu peuplés, sans surstockage et possédant des terres privées, se désertifient aussi gravement que n’importe quelle partie de l’Afrique, m’a amené à réaliser que nous devions chercher ailleurs. Je crois avoir trouvé cette cause sous-jacente dans la façon dont les gens à travers les âges ont réussi à améliorer nos vies – l’objectif principal du management.
Même si nous pensons qu’il existe de nombreuses façons de gérer et que les humains prennent des millions de décisions de gestion de plusieurs manières, même si elles sont dépouillées jusqu’à l’essentiel, comme éplucher les couches d’un oignon, même la gestion interdisciplinaire la plus sophistiquée repose sur un cadre de base. Découvrir l’existence de ce cadre m’a amené à comprendre que tous les gouvernements, agences de développement et ONG utilisent exactement ce même cadre lors de la formulation de politiques et de projets. Par souci de simplicité, j’appelle cela le cadre universel ou réductionniste.
IC. Cadre réductionniste universel utilisé par tous les gouvernements pour formuler des politiques
Les actions de gestion conscientes, par opposition aux instincts, qui traitent de tout problème lié à l’élaboration d’une politique sont menées en vue d’atteindre un objectif . Les seuls outils permettant de gérer l’environnement dans son ensemble qui sont pris en compte dans les politiques ou projets de tout gouvernement (ou agence de développement) relèvent des catégories de technologie , d’incendie ou de repos/conservation (de l’environnement). Au-delà de ces outils pour agir sur l’environnement, les gouvernements n’ont plus qu’une action possible : utiliser la technologie pour planter des arbres, des arbustes ou des herbes . Et toutes les actions visant à atteindre un objectif sont toujours basées sur un ou plusieurs facteurs parmi de nombreux facteurs, tels que l’expérience passée, l’opinion d’experts, les résultats de la recherche, l’opinion publique, le coût, le compromis, l’opportunité, les croyances culturelles, l’intuition, la pression des pairs, la peur, la propagande, coût, flux de trésorerie, rentabilité, etc.
Il n’y a aucune exception à l’utilisation de ce cadre simple dans la prise de décision consciente ; c’est ce qu’une simple famille d’éleveurs utilise quotidiennement, et c’est ce que les équipes scientifiques les plus sophistiquées utilisent également pour lutter contre la désertification, le changement climatique mondial ou l’exploration spatiale. Tous les gouvernements utilisent involontairement ce cadre, qui est, je crois, génétiquement ancré chez l’homme, lors de la formulation de politiques de gestion des ressources, ainsi que d’autres politiques.
IDENTIFIANT. Domaines où le cadre réductionniste universel est efficace.
Le cadre réductionniste s’est révélé efficace dans le développement de la technologie, depuis les outils de l’âge de pierre jusqu’aux machines sophistiquées utilisées pour les voyages spatiaux. Le succès stupéfiant de la technologie est considérable dans l’amélioration de la vie des gens grâce à la production alimentaire industrielle commerciale, aux services de santé, aux transports, à de nombreux dispositifs permettant d’économiser du travail et à des distractions divertissantes comme la télévision.
Même si nos succès technologiques améliorent généralement la vie des personnes riches, cela n’est pas vrai pour beaucoup de personnes, en particulier dans les zones rurales. Nos remarquables succès technologiques ne réussissent en réalité que si nous ignorons leurs effets à long terme sur notre environnement et notre société. Ces effets deviennent de plus en plus graves et menacent le bien-être futur de toutes les nations.
En science des systèmes, tous les domaines de réussite technologiques, qui sont en réalité tout ce que nous fabriquons plutôt que de gérer, sont décrits comme des systèmes durs et compliqués . En bref, cela signifie qu’ils sont conçus par des humains (une montre, un téléphone portable ou un ordinateur en sont de bons exemples) et qu’ils possèdent les caractéristiques suivantes :
Ils sont compliqués
Ils ne fonctionnent pas s’il manque des pièces
Ils possèdent des propriétés émergentes (ce qui signifie que toutes les pièces assemblées font ce qui a été conçu mais ne peuvent être réalisées par aucune des pièces séparément) : une montre vous indique l’heure, un téléphone vous permet de parler à quelqu’un.
Ils ne présentent pas de propriétés émergentes imprévues (ils font uniquement ce pour quoi ils sont conçus)
Lorsque des problèmes surviennent, ils sont relativement faciles à résoudre (appelés « bons problèmes »).
C’EST À DIRE. Des domaines où le cadre réductionniste universel donne moins de résultats.
Pour mieux comprendre les domaines de notre vie où le cadre réductionniste est moins efficace, nous devons nous tourner à nouveau vers la science des systèmes, qui reconnaît également les systèmes complexes (par exemple, les organisations humaines) et les systèmes naturels (par exemple, les plantes, les animaux, les sols, notre environnement). . Les systèmes doux et naturels, vous le reconnaîtrez, englobent tout ce que nous ne fabriquons pas mais que nous gérons. Les systèmes logiciels sont conçus par des humains. Les systèmes naturels ne le sont pas. Hormis cette différence, les systèmes doux et naturels ont les mêmes caractéristiques :
Ils impliquent des choses que nous gérons et ne doivent pas toujours impliquer la technologie.
Ils peuvent être compliqués, mais ils sont toujours complexes, dans la mesure où ils ont des propriétés émergentes mais aussi des propriétés imprévues ou inattendues (par exemple, une organisation fera ce qu’elle était prévue de faire mais fera également des choses imprévues ou attendues).
Ils s’auto-renouvellent ou s’auto-organisent
Ils travaillent avec des pièces manquantes
Lorsque des problèmes surviennent, ils sont extrêmement difficiles à résoudre (appelés problèmes épineux).
C’est dans ces domaines de notre vie – avec tout ce que nous gérons – qui impliquent la complexité des systèmes souples et naturels qui, en utilisant comme tous le font, le cadre réductionniste, nous nous heurtons à des problèmes et des conflits sans cesse croissants à travers le monde. Nombreux sont les succès apparemment mineurs, mais si on les considère à grande échelle et avec honnêteté, nous perdons du terrain à mesure que la population augmente et que la désertification et le changement climatique mondial s’accélèrent. Lorsque des nations entières, y compris les États-Unis, exportent davantage de terres érodées que de céréales, de viande, de bois, de produits commerciaux et militaires, elles se dégradent rapidement. L’estimation récente de 4 tonnes de sol en érosion qui s’écoulent chaque année dans les rivières du monde pour chaque être humain vivant nous révèle l’ampleur mondiale du problème de la mauvaise gestion des ressources.
Cette digression dans la science des systèmes, aussi brève soit-elle, est essentielle pour comprendre les propriétés émergentes imprévues/inattendues des bureaucraties de la fonction publique héritées du Zimbabwe et d’autres pays africains qui rendent la bonne gouvernance impossible, comme je l’explique dans la section III.
SI. Pourquoi les politiques actuelles de gestion des ressources sont judicieuses.
Par souci de brièveté, je ne mentionne que les deux principaux défauts du cadre réductionniste qui conduisent généralement à des politiques et à des projets malsains qui tentent de résoudre les problèmes de société ou de ressources naturelles.
Lacunes des actions entreprises dans un contexte ou une raison trop simple pour un réseau d’une complexité inévitable.
Réduire le réseau inévitable de complexité sociale/culturelle, économique et environnementale définit une gestion réductionniste. Une telle gestion défie la réalité d’un monde fonctionnant de manière holistique et constitue la raison la plus fondamentale de tant de nos maux. Bien que les objectifs et les buts des politiques ou des projets soient souvent atteints à court terme, nous subissons par la suite des conséquences imprévues et le besoin de solutions de plus en plus nombreuses. Des livres entiers ont été écrits sur ce problème, sur lequel je n’ai pas besoin d’insister.
Aucun outil permettant de prévenir ou d’inverser la désertification sur la plupart des terres du monde. La société, et donc les scientifiques, ont toujours cru que la désertification était causée par le surpâturage du bétail et qu’elle pouvait donc être inversée en mettant fin au surpâturage en réduisant ou en supprimant le bétail. Des milliers d’articles publiés, évalués par des pairs, attestent de telles croyances. En réalité, la désertification se produit lorsque les précipitations disponibles deviennent moins efficaces. La pluie efficace est la pluie qui tombe et pénètre dans le sol, puis ne quitte le sol que par deux voies. Il sort en transpirant à travers les plantes ou en s’écoulant dans le sol jusqu’aux rivières, zones humides ou réservoirs souterrains. Les précipitations non efficaces sont celles qui tombent et traversent le sol, provoquant des crues soudaines, ou bien elles pénètrent dans le sol et le quittent ensuite en s’évaporant des surfaces nues du sol. À mesure que les précipitations deviennent moins efficaces, la fréquence et la gravité des inondations et des sécheresses augmentent, sans réel changement dans les précipitations. Le fait que la désertification se produise ou non dépend du pourcentage de sol nu entre les plantes sur des milliards d’hectares de terre.
Dans tous les environnements du monde, il n’y a que deux choses que font les humains qui conduisent à des milliards d’hectares de sol en grande partie nu entre les plantes, comme nous le voyons partout au Zimbabwe et dans les pays voisins – en fait environ les deux tiers de la superficie mondiale. Ces deux choses sont une gestion qui aboutit à ce que trop peu de gros animaux errent autour des plantes en surpâturage tout en reposant excessivement la terre et/ou en utilisant le feu. Rien d’autre ne conduit à des millions d’hectares de sol en grande partie nu entre les plantes dans n’importe quel environnement terrestre du monde. Malheureusement, ces deux pratiques sont courantes chez les scientifiques et les gouvernements ainsi que dans la société dans son ensemble et la propagation mondiale de la désertification n’est donc pas un mystère.
Lorsque l’on examine les « outils » dont disposent les humains pour gérer notre environnement dans son ensemble, nous remarquons qu’il y en a deux (le feu et le repos/conservation) qui favorisent la désertification sur environ les deux tiers de la surface terrestre du monde soumise à des précipitations saisonnières et/ou irrégulières. Et nous notons qu’il n’existe aucun outil capable d’inverser la désertification, ce qui ne peut être réalisé en utilisant aucune technologie imaginable, ni en utilisant la technologie pour planter des arbres à l’échelle requise et là où les précipitations sont insuffisantes pour maintenir une couverture complète du sol sous les arbres. Ainsi, cela aurait été un miracle si les terres ne se dégradaient pas dans une grande partie du monde et si les déserts n’avaient pas progressé au cours de l’histoire. L’opinion générale est qu’il existe un grand nombre d’« outils » et donc d’options disponibles pour résoudre les problèmes environnementaux. En réalité, formez-vous à n’importe quelle profession dans n’importe quelle université dans le monde et, sans le savoir, vous ne serez formé qu’à utiliser la technologie, le feu, le repos/conservation ou la technologie pour planter des arbres et d’autres plantes afin de gérer notre environnement dans son ensemble.
Ainsi, pour des raisons à la fois de gestion et de science, il est désormais compréhensible qu’aucun gouvernement, ni aucune agence de développement, n’ait jusqu’à présent été capable de produire des politiques qui soient simultanément économiquement, socialement et écologiquement rationnelles à court et à long terme dans la majeure partie du monde. y compris le Zimbabwe, les pays voisins et même les États-Unis.
Remarque : Il existe des cas mineurs dans des environnements perpétuellement humides où il est théoriquement possible pour les gouvernements de créer des politiques holistiques solides, mais dans la pratique, cela est rare, non pas à cause d’un manque d’outil, mais à cause d’une gestion réductionniste.
IG. Cadre de gestion holistique.
Ma quête pour comprendre la désertification en cours en Zambie et au Zimbabwe à partir des années 1950 m’a amené à développer un certain nombre d’idées que j’ai pu tester en pratique avec des gestionnaires de terres sur quatre continents. Cette quête m’a également amené à examiner le travail d’autres scientifiques, principalement au Zimbabwe, en Afrique du Sud, en France et aux États-Unis, et à développer progressivement un cadre de gestion de base amélioré. Ce nouveau cadre de gestion est décrit dans Holistic Management : A Commonsense Revolution to Restore Our Environment , (Island Press) 2016.
En bref, le cadre de gestion holistique améliore le cadre réductionniste avec quelques ajouts :
Un contexte holistique déterminé par les personnes qui gèrent ou élaborent des politiques qui relient ce que les gens apprécient le plus dans la vie à leur soutien à la vie.
Reconnaissance que toute activité humaine dépend de la santé de notre environnement – à savoir quatre processus qui régissent le cycle de l’eau, le cycle des nutriments, la diversité et la stabilité des communautés biologiques et le flux total de lumière solaire pour soutenir toute vie, y compris les humains.
L’ajout de deux outils qui rendent possible l’inversion de la désertification dans les environnements de précipitations saisonnières du monde : le pâturage et l’impact des grands herbivores tels que le bétail.
Un ensemble de questions de vérification du contexte pour garantir que les actions sont conformes au contexte holistique et donc socialement/culturellement, économiquement et environnementalement saines à court et à long terme, menant vers les personnes futures.
Comme toutes les nouvelles innovations, la gestion holistique n’a pas été acceptée ou adoptée rapidement, mais aujourd’hui le livre référencé ci-dessus est utilisé dans de nombreuses universités, et il existe un réseau mondial de centres de gestion holistique dirigés et gérés localement sur six continents, largement calqués sur le premier modèle. un tel hub au Zimbabwe.
IH. Le contexte holistique global unique pour guider la gestion et la politique.
La différence la plus essentielle entre la gestion réductionniste et la gestion holistique est que, alors que dans la première nous avons de nombreux besoins, désirs ou problèmes différents à résoudre et que ceux-ci constituent la raison ou le contexte de nos actions, avec la gestion holistique, nous avons maintenant un seul contexte holistique global. pour guider toutes les actions lorsque nous répondons à nos besoins, nos désirs ou résolvons des problèmes.
L’une des plus grandes conséquences involontaires et méconnues de la gestion réductionniste, y compris l’élaboration de politiques, est qu’elle conduit généralement à des conflits et même à des guerres occasionnelles. En effet, sans un contexte unificateur pour les actions, les gens promeuvent ou soutiennent des actions qui aboutissent à des conflits et à la violence. Lorsque des personnes, même de cultures et de croyances différentes, développent d’abord un contexte holistique avec un accord total, ce qui est essentiel, les actions ultérieures dans ce contexte sont généralement acceptables pour tous.
Même si cela peut paraître utopique, c’est pourtant réel. En travaillant avec de nombreuses personnes et cultures, j’ai toujours constaté que les gens voulaient essentiellement la même qualité de vie. Et là où j’ai travaillé avec des gens en conflit violent qui me criaient dessus et me disaient qu’il était impossible d’obtenir un accord au sein de leur communauté, je les ai amenés à accepter de travailler avec moi pendant seulement une heure et à me prouver le contraire. Une heure plus tard, ils étaient totalement d’accord sur un contexte holistique pour leur vie.
Le cadre de gestion holistique peut être utilisé dans n’importe quelle situation, de la vie d’une seule personne à celle d’un ménage, d’une entreprise ou d’une nation – tout comme le cadre réductionniste est utilisé universellement dans n’importe quelle situation aujourd’hui.
Même si le contexte global doit être développé par les décideurs impliqués dans la gestion, cela n’est pas toujours pratique au niveau national ou international dans les premières étapes de l’élaboration des politiques. Lorsque cela se produit, les personnes à l’origine de la politique utilisent un contexte holistique générique pour guider le processus jusqu’au point où un véritable contexte holistique peut être développé par les personnes appropriées. Dans les situations impliquant une politique potentiellement controversée, un véritable contexte holistique est nécessaire pour éviter les conflits et unir les gens. Un contexte holistique générique représente ce que veulent la plupart des gens et est donc remarquablement similaire à tout contexte holistique réel final – la différence réside dans le sentiment d’appropriation profonde dans ce contexte pour guider la gestion.
Pour me permettre de m’assurer que les suggestions (actions) que je fais dans ce document sont susceptibles de conduire à un Zimbabwe meilleur pour tous mes concitoyens, je dois moi aussi éviter que ma raison ou mon contexte soit réduit au problème de la mauvaise gouvernance. d’aujourd’hui. J’ai donc dû d’abord développer un contexte national holistique générique pour guider mes réflexions et mes écrits sur la manière dont nous pourrions réaliser ce que tous désirent. Parvenir à une bonne gouvernance est un objectif. Beaucoup ont essayé tout au long de l’histoire et ont échoué simplement parce qu’un objectif atteint dans le contexte de la résolution d’un problème avait peu de chances de réussir et était susceptible d’entraîner des conséquences inattendues.
Voici le contexte holistique générique national que j’ai utilisé dans la rédaction de cet article pour guider ma nation.
Qualité de vie : (Ce que nous voulons que notre vie soit basé sur ce que nous apprécions le plus dans la vie)
Nous voulons vivre en paix et en harmonie avec nous-mêmes et avec les pays voisins. Nous voulons la prospérité, la sécurité physique et financière. Bonne éducation pour nos enfants à tous les niveaux. Liberté de poursuivre nos propres croyances culturelles, religieuses et spirituelles. De bons logements et commodités dans nos villes. Des familles stables bénéficiant d’une sécurité alimentaire adéquate, d’aliments sûrs, nutritifs et sains et d’eau potable en abondance. Vivre en équilibre avec nos ressources avec un équilibre entre les populations urbaines et rurales afin que tous puissent vivre dans la paix et la prospérité. Accès juste et équitable aux ressources. Liberté de toute bigoterie raciale, tribale, sexuelle ou de tout autre sectarisme, avec une justice accessible et abordable pour tous. Fierté de nous-mêmes, de notre pays et de ses réalisations. Respect international. Jouer notre rôle en tant que nation dans les affaires internationales en tant qu’égaux respectés.
Base de ressources future : (Comment nous devons nous comporter et à quoi doit ressembler notre terre dans mille ans pour soutenir les générations successives vivant de telles vies) Comportement : En tant que nation, nous devons être honnêtes, justes, amicaux et ouverts avec une bonne attitude envers les étrangers et les autres nations.
Terre : Nos sols sur les terres cultivées et les parcours doivent être recouverts et régénérés en permanence d’une biodiversité et d’un volume de vie élevés, retenant l’eau et convertissant l’énergie solaire en richesse et en vie. Les rivières doivent également fonctionner en permanence avec des eaux souterraines abondantes.
II. Ampleur d’une politique malsaine révélée par un cadre holistique.
Comme mentionné, le cadre holistique a été développé à l’origine pour comprendre et inverser la désertification de manière pratique et peu coûteuse. Ce n’est qu’après son développement que nous avons découvert que le cadre holistique pouvait être utilisé d’une manière impossible avec une gestion réductionniste. En particulier, le cadre holistique peut être utilisé pour analyser les politiques et les projets de développement avant leur mise en œuvre, ou pour concevoir des politiques et des projets globalement solides, plus susceptibles de réussir.
Par exemple, au début des années 1980, quelque 2 000 scientifiques issus de nombreuses disciplines impliquées dans la gestion des ressources, provenant d’agences de gestion des terres du gouvernement américain, d’universités (agricoles) concédant des terres, de la Banque mondiale, de l’USAID et du US Fish& Wildlife Service, ont été formés à l’utilisation de la méthode holistique. cadre. Dans le cadre de cette formation, ils devaient apporter des exemples de leurs propres politiques à analyser. Toutes ces politiques, sans exception, se sont révélées défectueuses et sans aucune chance de succès. L’un de ces groupes en formation a déclaré à l’unanimité qu’ « ils pouvaient désormais reconnaître qu’une mauvaise gestion des ressources était universelle aux États-Unis. « Une formation similaire en Inde, au Lesotho et au Zimbabwe a abouti à des résultats similaires, bien qu’avec des échantillons plus petits.
Malgré la bonne intention des études d’impact environnemental exigées par de nombreuses autorités avant l’acceptation d’une politique ou d’un projet, aucune EIE, car toutes utilisent le cadre réductionniste universel, n’a pu détecter les failles politiques. Le problème est, je crois, universel, mais n’importe quel gouvernement pourrait le résoudre rapidement et à peu de frais grâce à la formation, comme commence à le faire le Savory Institute de Boulder, Colorado.
Ghandi, lorsqu’on lui a demandé ce qu’il pensait de la civilisation occidentale, aurait répondu : « C’est une bonne idée ». Ce n’est que lorsque les gouvernements seront capables d’élaborer des politiques ou des projets globalement solides que la bonne gouvernance deviendra plus qu’un simple idéal ou une idée.
Cependant, aussi importante que soit une politique saine, il existe d’autres conditions à remplir avant que tous se sentent en sécurité et bien gouvernés.
Section II. Le problème des partis politiques
Sachant que les systèmes à parti unique aboutissent inévitablement à des abus de pouvoir, à des dictatures (militaires ou autres) et à la violence, la croyance largement répandue en une démocratie multipartite est compréhensible, tout comme le désir du monde occidental d’imposer de telles croyances aux démocraties naissantes en Afrique. . Cependant, la croyance selon laquelle les partis politiques eux-mêmes sont essentiels au gouvernement démocratique bloque la pensée créative et empêche, selon moi, la réalisation d’une bonne gouvernance. Je ne suis pas le premier à voir les dangers des partis politiques pour la démocratie et la stabilité des nations. George Washington, qui a refusé de briguer un autre mandat à la présidence des États-Unis, dans son discours d’adieu à sa nation en septembre 1796, a transmis cet avertissement à propos des partis politiques :
» Sans espérer une extrémité de ce genre (qui ne devrait cependant pas être entièrement hors de vue), les méfaits communs et continuels de l’esprit de parti suffisent pour faire de l’intérêt et du devoir d’un peuple sage de décourager et de retenez-le.
Il [le système des partis] sert toujours à distraire les conseils publics et à affaiblir l’administration publique. Il agite la Communauté avec des jalousies mal fondées et de fausses alarmes ; attise l’animosité des uns contre les autres, fomente parfois des émeutes et des insurrections. Cela ouvre la porte à l’influence étrangère et à la corruption, qui trouvent un accès facilité au gouvernement lui-même par le canal des passions partisanes. Ainsi, la politique et la volonté d’un pays sont soumises à la politique et à la volonté d’un autre.
Même si l’anglais est d’un autre âge, le message est clair. Les dirigeants africains feraient bien de tenir compte de cet avertissement sur le danger d’autoriser les partis politiques, de la part de George Washington, qui a conduit son pays à l’indépendance de la Grande-Bretagne et qui recherchait la bonne gouvernance et non le pouvoir.
Sur la base de ma participation à des partis politiques en tant que candidat et/ou leader, et sur la base de mes observations de la scène politique dans de nombreux pays, j’ai conclu que l’existence de partis politiques constitue le plus grand obstacle au gouvernement démocratique.
Aujourd’hui, dans de nombreux pays, les gens manifestent leur dégoût, leur frustration et leur sentiment de désespoir en ne prenant pas la peine de voter. J’entends souvent les gens dire : « À quoi ça sert de voter, ça ne fait aucune différence ». Personnellement, je ressens cela aussi. Privé de mon vote dans mon propre pays, j’ai le droit de voter aux États-Unis, mais je ne vois souvent pas d’autre intérêt que d’essayer de minimiser les dommages causés aux Américains en votant pour le parti le moins dommageable, en termes de politiques promues par ce parti. Récemment, nous avons vu le mécontentement à l’égard des politiques gouvernementales conduire au Brexit en Grande-Bretagne et au phénomène Trump aux États-Unis – sans aucune possibilité qu’une telle frustration exprimée ne conduise à autre chose que les mêmes vieilles politiques destructrices avec des conséquences inattendues.
Pour les raisons expliquées ci-dessous, le système des partis ne fonctionne tout simplement pas, en particulier en Afrique, et ne pourra probablement jamais garantir une bonne gouvernance dans aucun pays. Dans la section III, je présente des suggestions sur la manière dont une démocratie sans parti pourrait former un gouvernement d’une manière plus véritablement démocratique.
II-A. Croyance fondamentale requise pour que le système de partis fonctionne.
Pour qu’un comportement politique pacifique et civilisé puisse avoir lieu pendant les élections, il est essentiel que les électeurs et les candidats croient profondément à l’idée d’une « opposition loyale ». Ce concept, né après des siècles de luttes et de conflits en Europe, est une croyance que les peuples d’Afrique et d’ailleurs ne partagent tout simplement pas. La croyance est plutôt que quel que soit le parti qui accèdera au pouvoir, il récoltera les bénéfices et ferait mieux de rester au pouvoir à tout prix, car il ne bénéficiera plus jamais d’un accès aussi facile à la richesse.
Généralement, le parti qui accède au pouvoir lors des premières élections africaines post-indépendance fera de grands efforts pour ne pas abandonner démocratiquement le pouvoir. Lorsque les partis nationalistes africains se sont battus pour l’indépendance sous la bannière de la démocratie avec le slogan « Un homme, une voix », de nombreuses personnes comme moi ont soutenu leurs aspirations dans nos cœurs. Cependant, dans notre esprit, nous savions que cela signifiait probablement « un homme, une voix, une fois ». Et l’histoire a montré que c’était généralement le cas. Une fois que le peuple a eu son premier et unique vote, qui a porté le parti de son choix au pouvoir, ce fut la fin de tout semblant de démocratie pour les années à venir et c’est toujours le cas aujourd’hui au Zimbabwe après près de quarante ans.
Trop souvent, ce n’est qu’après beaucoup de violence et de souffrance que le parti au pouvoir est remplacé par un autre et le cycle se poursuit avec des partis successifs faisant tout ce qu’ils peuvent pour rester au pouvoir en refusant aux citoyens tout autre choix démocratique. Tous les partis politiques, lorsqu’ils cherchent le pouvoir, prétendent soutenir la démocratie. Cependant, les partis, comme les citoyens, doivent être jugés non pas sur leurs paroles mais sur leur comportement. Ce comportement des partis politiques ne devrait surprendre personne là où les gens ne croient pas en une opposition loyale. Ce serait franchement un comportement anormal s’il n’y avait pas de manipulation et de violence à des degrés divers de la part d’un parti politique au pouvoir.
Certains États démocratiques multipartites, comme la Grande-Bretagne, affichent leur croyance culturelle en une opposition loyale à travers le comportement de leur gouvernement et de leur électorat. Chaque parti en lice n’a pas besoin de distribuer des T-shirts et des machettes et sait que s’il ne gagne pas, il ne sera pas interdit et ses membres ne seront pas battus, tués ou torturés. Il formera une opposition efficace et aura de bonnes chances de gagner aux prochaines élections. Dans ces pays, le parti au pouvoir permet aux autres partis de se présenter aux élections et à l’électorat de déterminer le résultat, comme l’a fait le gouvernement britannique lorsque le Parti communiste s’est présenté aux élections. D’autres pays, comme les États-Unis, soutiennent du bout des lèvres le concept d’une opposition loyale, comme en témoigne l’interdiction du Parti communiste apparu dans les années 1950. En Amérique, contrairement à la Grande-Bretagne, l’électorat n’a pas été autorisé à déterminer l’issue du scrutin. Le gouvernement a interdit le Parti communiste et s’est lancé dans d’effroyables chasses aux sorcières, détruisant la vie de nombreux citoyens.
II-B. Les partis donnent la priorité à leur nation seulement en temps de tragédie ou de guerre
Les gouvernements fondés sur un système de partis quel qu’il soit ne se rapprochent de l’unité nationale que lorsque les partis politiques collaborent dans l’intérêt national sous une menace extérieure, comme en cas de guerre.
Toutefois, la collaboration en temps de guerre reste loin de répondre aux exigences d’une bonne gouvernance et, une fois la guerre terminée, les parties se retrouvent inévitablement à nouveau engagées dans des luttes de pouvoir au détriment de la nation.
II-C. Loyauté des forces armées à faire la fête
En Afrique et ailleurs, les partis au pouvoir manipulent régulièrement les forces armées de leur pays en encourageant, voire en renforçant, la loyauté envers le parti au-dessus de la nation. En tant que soldat et homme politique, j’ai vécu et combattu pendant la longue guerre d’indépendance du Zimbabwe. Moi-même et une poignée d’officiers de l’armée étions pleinement conscients que la guerre aurait pu être évitée si nos généraux avaient respecté le serment d’allégeance que nous avons prêté lors de notre entrée en fonction. Notre serment d’allégeance était envers notre nation et non envers un parti politique. En prenant le pouvoir, le parti du Front rhodésien (RF) dirigé par Ian Smith a rapidement remplacé les généraux récalcitrants. Les généraux nouvellement nommés, favorables aux politiques raciales de la Fédération de Russie, alignèrent bientôt les forces armées sur le parti politique raciste de Smith. Presque immédiatement, le parti a pris le contrôle des médias et du système judiciaire et, du jour au lendemain, toute critique de Smith ou de son parti a été interprétée comme une déloyauté envers la nation. Smith n’a pas mis longtemps à modifier la constitution, créant 50 sièges réservés aux Blancs et 16 sièges noirs « sur les bancs latéraux », privant ainsi de fait la plupart des Zimbabwéens du droit de vote. Lorsque, faisant preuve de bon sens, j’ai dit publiquement que Smith devait parler à Nkomo et Mugabe, Smith et une bande de fidèles du parti ont demandé que je sois jugé pour trahison. La guerre prolongée et les pertes en vies humaines qui ont suivi étaient inévitables, tout comme mon éventuel exil.
Que les forces armées du Zimbabwe, après l’indépendance, se soient alignées sur le parti de Mugabe plutôt que sur la nation du Zimbabwe n’était en rien inhabituel en Afrique. Il n’était pas non plus inhabituel que des officiers faisant preuve d’une allégeance aveugle au parti au-dessus de la nation soient généreusement récompensés par des fermes et des entreprises confisquées aux citoyens zimbabwéens. Ces pratiques ne se produiraient dans aucune nation véritablement démocratique ou sans parti.
II-D. Les politiques prédéterminées des partis ne peuvent assurer une bonne gouvernance
Si je n’avais pas traversé le Parlement et formé un parti politique de toutes pièces une fois que Smith avait effectivement détruit tout semblant de démocratie dans notre pays, je n’aurais jamais compris comment les programmes des partis influencent les positions politiques.
Les partis cherchent à se faire élire sur la base de leur programme déclaré, qui exprime les convictions générales des personnes qui soutiennent ce parti et, en général, au moins les grandes lignes des politiques que le parti poursuivra s’il est élu. Cependant, comme expliqué dans la section I, les politiques prédéterminées ne répondent pas à toute la complexité inhérente à la complexité sociale, environnementale et économique d’un pays. C’est encore plus vrai lorsque les programmes électoraux des partis répondent à l’attrait émotionnel et économique à court terme des électeurs et sont généralement réduits à des slogans émotifs. Il n’est donc pas surprenant que le parti vainqueur, représentant les convictions de ses partisans, laisse mécontents ceux qui ont soutenu les autres partis et font de leur mieux pour s’opposer à de telles politiques.
Lorsque, plus tard, les inévitables complications et lacunes politiques provoquent colère et frustration, les citoyens commencent à compter les jours jusqu’aux prochaines élections et à aiguiser leurs machettes. Si un autre parti prend le pouvoir, le cycle se répète comme il l’a fait au cours des siècles de quête du pouvoir par les partis.
II-E. Les partis dont les politiques sont basées sur «…. Les «ismes» ne parviennent pas à assurer une bonne gouvernance.
Les partis politiques du monde entier ont pour fondement politique fondamental l’essai de toutes sortes de «…ismes» : le capitalisme , le communisme , le socialisme , le racisme , le tribalisme , le copinage et le corporatisme qui émerge actuellement. Aucun parti fondé sur un tel «…isme» ne peut assurer une bonne gouvernance, pour les raisons exposées dans la section I, et aussi parce qu’il y aura toujours une proportion de l’électorat qui ne partage pas les croyances, l’idéologie ou le dogme de ces partis et donc ne se sent ni en sécurité ni bien gouverné.
II-F. Les partis fondés sur des personnalités ou sur la religion ne peuvent assurer une bonne gouvernance.
Les partis fondés sur des personnalités ou sur la religion ont tendance à conduire à une dictature ou à une tyrannie qui se termine par un renversement violent à un moment donné, soit au niveau interne, soit par une invasion lorsque leurs politiques menacent les voisins d’une autre conviction. Il est courant que des dirigeants de haut niveau, ou des partis accrochés au pouvoir, créent des conflits, voire des guerres ou d’autres distractions, afin d’éviter de régler des problèmes intérieurs. Même si une politique foncière était attendue depuis longtemps au Zimbabwe, le vol soudain de fermes pour les redistribuer aux partisans du parti par le gouvernement du président Mugabe était sans aucun doute une telle tactique de diversion à une époque où le chômage, la demande d’emploi et d’autres mécontentements avaient conduit à la formation d’une véritable politique foncière. parti d’opposition. Comme la plupart des tactiques de distraction des partis politiques, celle-ci s’est avérée coûteuse. Je reviendrai sur la politique foncière, encore indispensable, et qui n’existe toujours pas.
Les partis fondés sur des croyances religieuses, s’ils provoquent de telles distractions contre les autres, représentent un grand danger non seulement pour eux-mêmes mais aussi pour les autres. Tout parti représentant un groupe religieux présente un problème supplémentaire dans la mesure où les religions organisées d’aujourd’hui sont elles-mêmes divisées et souvent en conflit. Par exemple, il existe bien plus de 1 000 branches du christianisme qui posent à elles seules un problème à tout parti fondé sur le christianisme. Quelle que soit la branche qui prendra le pouvoir, d’autres branches résisteront inévitablement, comme l’ont démontré des années de conflit entre catholiques et protestants en Irlande. Les conflits actuels au sein de la religion islamique sont évidents pour tous et destructeurs pour de nombreuses nations, alors que différents éléments recherchent le pouvoir politique religieux en utilisant les gilets suicides comme une avancée technologique par rapport aux machettes.
En général, toutes les religions organisées présentent le même problème car, en théorie, même si une religion était absolument unie, tous les citoyens d’une nation ne sont pas tous d’une même foi. Un exemple est celui du Bhoutan, qui lutte pour un système démocratique et la mesure des progrès par le bonheur national brut pour la plupart des Bhoutanais, qui sont bouddhistes, tandis que la minorité chrétienne serait supprimée.
II-G. Partis politiques sujets à la corruption.
La corruption prend de nombreuses formes, l’une d’elles étant l’influence indue exercée sur un gouvernement pour qu’il suive des politiques davantage dans l’intérêt de groupes d’intérêts spéciaux ou d’entreprises que dans celui des personnes qui ont élu le parti au pouvoir. Comment expliquer autrement que les gouvernements entrent en guerre pour protéger les intérêts des entreprises en conflit direct avec les intérêts de leurs citoyens ?
Je me suis souvent demandé pourquoi les Américains qui sont si gentils et généreux (probablement la nation la plus généreuse de tous les temps) ne sont pas aimés et appréciés dans le monde entier ? Pour répondre à cette question et dans quelle mesure elle est liée à la facilité avec laquelle les partis politiques succombent à la corruption, nous devons rappeler l’avertissement de George Washington sur les dangers des partis politiques donnés à son peuple pour obtenir son indépendance.
Même si j’utilise ici l’exemple des États-Unis, les Américains n’ont pas le monopole de la mauvaise gouvernance. La colère internationale contre les Américains n’est pas difficile à comprendre si l’on considère la politique étrangère américaine sous l’un ou l’autre parti politique. Le général Smedley Butler, l’un des chefs expéditionnaires des Marines les plus célèbres, à l’heure de sa retraite de l’armée américaine, a déclaré ceci : « J’ai passé 33 ans et 4 mois en service militaire actif… Et pendant cette période, j’ai passé la plupart de mon temps en tant qu’homme musclé de haut niveau. pour les grandes entreprises, pour Wall Street et les banquiers. Bref j’étais un racketteur, un gangster du capitalisme. Ainsi, j’ai contribué à rendre le Mexique et surtout Tampico sûrs pour les intérêts pétroliers américains en 1914. J’ai contribué à faire d’Haïti et de Cuba des endroits décents où les garçons de la National City Bank pouvaient collecter des revenus. J’ai contribué au viol d’une demi-douzaine de républiques d’Amérique centrale pour le compte de bénéfice de Wall Street. J’ai aidé à purifier le Nicaragua pour la maison bancaire internationale Brown Brothers en 1902-1912. En 1916, j’ai fait la lumière sur les intérêts américains du sucre en République Dominicaine. En 1903, j’ai contribué à faire du Honduras un pays favorable aux entreprises fruitières américaines. En Chine, en 1927, j’ai contribué à ce que la Standard Oil continue son chemin sans être inquiétée.»
Les gouvernements élaborant une politique étrangère pour servir les intérêts des entreprises plutôt que ceux de leurs citoyens sont devenus plus, et non moins, omniprésents depuis l’époque du général Butler et je souligne à maintes reprises qu’il ne s’agit pas d’un monopole américain.
Ainsi, les gens bons et amicaux dans de nombreux pays sont enclins à se retrouver dans des conflits et des guerres fondés, non pas sur les intérêts ou les souhaits de leur peuple, mais sur les intérêts des entreprises qui corrompent la politique des partis et la politique étrangère. La Grande-Bretagne a finalement dû maîtriser la Compagnie des Indes orientales, mais pas avant que l’armée de la compagnie ne soit plus nombreuse que celle de la Grande-Bretagne et que la compagnie ait ses propres juges et prononce même des condamnations à mort. La Boston Tea Party, où les gens jetaient le thé de la Compagnie des Indes orientales depuis leurs navires dans le port, fut l’une des premières étapes menant à la Révolution américaine et à l’indépendance.
Les pères fondateurs de l’Amérique ont tenté de garantir que les intérêts des citoyens prévaudraient dans une république populaire indépendante. Souvenez-vous de l’avertissement de Washington à propos des partis politiques : « les méfaits communs et continus de l’esprit de parti suffisent pour qu’il soit dans l’intérêt et le devoir d’un peuple sage de le décourager et de le restreindre. » Ignorant ses conseils, le système des partis a été adopté par les Américains. Le développement des partis, malgré de vaillantes tentatives pour réduire le pouvoir excessif en ayant trois branches de gouvernement, a permis la manipulation des entreprises, avec le consentement volontaire des partis politiques et du système judiciaire américain, au fil des années pour conduire au pouvoir des entreprises à diriger pratiquement l’Amérique, quel que soit le parti. est au pouvoir. Comme je le dis à mes amis américains, vous n’avez pas de démocratie telle que je la connais parce que votre seul choix est de voter pour la gauche ou pour la droite du même parti corporatif.
De nombreuses entreprises opèrent légitimement dans l’intérêt public comme prévu, mais comme pour le pouvoir politique, trop de richesse et de pouvoir corrompt. Il existe aujourd’hui une crainte légitime que les sociétés transnationales, dotées de budgets supérieurs à ceux de nombreux pays et n’ayant de comptes à rendre à aucun électorat, prennent le relais là où le colonialisme s’est arrêté. De plus, ces sociétés sont assistées par les gouvernements de nations puissantes et les agences internationales créées par eux, comme la Banque mondiale, le FMI et d’autres. La menace que représente cette nouvelle forme de colonialisme pour les nations africaines est en effet grave, car la Chine et d’autres pays servent leurs propres intérêts par l’intermédiaire des dirigeants des partis politiques africains. Les pays africains, tout comme les États-Unis, n’ont pas tenu compte de l’avertissement de George Washington concernant le système des partis politiques : « Il ouvre la porte à l’influence étrangère et à la corruption, qui trouvent un accès facilité au gouvernement lui-même par le canal des passions partisanes. Ainsi, la politique et la volonté d’un pays sont soumises à la politique et à la volonté d’un autre.
De nombreuses preuves suggèrent que les partis politiques sont si enclins à la corruption sous de nombreuses formes qu’ils font peu pour l’empêcher. Même si ce n’est pas toujours le cas, il est assez courant de penser que les gouvernements seraient moins corrompus sans les partis politiques.
Si les gouvernements voulaient réellement prévenir la corruption, en particulier au niveau gouvernemental, ils pourraient le faire. Un sondage auprès des citoyens aux États-Unis, en Grande-Bretagne, en France, en Italie et en Russie indiquerait presque certainement que la majorité n’est pas favorable au commerce international des armes. Malgré ce manque de soutien citoyen, le commerce international des armes reste l’une des plus grandes entreprises au monde, dominé par les États-Unis, la Grande-Bretagne, la France, l’Italie et la Russie et qui entretient des conflits sans fin, des souffrances et des massacres de personnes et d’animaux sauvages. Au moment où j’écrivais la première version de cet article, le gouvernement britannique était impliqué dans un énorme scandale très médiatisé concernant le trafic d’armes avec la famille royale saoudienne.
Le fait que les partis politiques soient généralement à l’avant-garde de la corruption en Afrique ne devrait pas surprendre. La principale différence entre la corruption de nos partis africains et celle des pays mentionnés réside uniquement dans le niveau de corruption officielle flagrante et le manque de sophistication. Ce n’est un secret pour personne que de nombreux hommes politiques et responsables de partis africains sont devenus, depuis l’indépendance, incroyablement riches tandis que leurs citoyens ont été contraints d’émigrer ou sont tombés dans la pauvreté et le désespoir. Grâce au système de partis politiques, le Zimbabwe a connu la mort de milliers de citoyens et l’appauvrissement d’une nation autrefois riche. Ce n’est qu’en interdisant les partis politiques afin de garantir une bonne gouvernance que de tels abus de la part des partis politiques pourront être évités.
II-H. Les partis politiques se concentrent davantage sur le maintien du pouvoir que sur la gouvernance.
Plutôt que de se concentrer à long terme sur la gouvernance, les partis politiques font inévitablement preuve d’une obsession à court terme de conserver le pouvoir lors des prochaines élections. La bonne gouvernance exige une planification et une continuité à long terme, ce qui n’est pas le point fort des partis confrontés à des élections toutes les quelques années. La réflexion à long terme n’est pas non plus le point fort des entreprises qui contrôlent ou influencent les partis politiques. Après tout, les PDG des sociétés publiques sont légalement tenus de fournir des rapports et des bénéfices trimestriels aux actionnaires et ne sont responsables devant l’électorat d’aucun pays.
II-I. Manipulation du parti et contrôle du système judiciaire et des médias
Trop souvent, les partis au pouvoir nomment des juges favorables à leurs convictions, bafouant ainsi l’idée d’un système judiciaire véritablement impartial et indépendant, essentiel à la bonne gouvernance et à la justice.
Il est rare que les partis obsédés par le maintien du pouvoir se contentent d’influencer le système judiciaire. Trop souvent, ils manipulent également les médias et interdisent généralement les organisations médiatiques indépendantes. Même dans les pays où les politiciens ne peuvent pas s’en sortir en contrôlant les médias, la collusion avec les entreprises qui contrôlent les médias est à peine déguisée. J’ai grandi pendant la Seconde Guerre mondiale en croyant à l’indépendance et à l’impartialité de la BBC, mais j’ai ensuite été témoin des tentatives de Margaret Thatcher de contrôler la BBC pendant la guerre des Malouines. Heureusement, la démocratie britannique était suffisamment mûre pour ne pas s’en tirer à bon compte. Le pouvoir judiciaire et les médias sont des cibles particulières, car tout parti qui les contrôle ou les influence suffisamment est presque assuré de réussir aux élections. Il en est ainsi parce que l’égalité, ou l’inégalité, des règles du jeu lors des élections n’est pas déterminée le jour du scrutin mais au fil des mois et des années précédant les élections.
Nous, les Zimbabwéens, aurions dû retenir cette leçon. La plupart des gens sont tellement insensibles aux effets quotidiens du contrôle des partis sur les médias qu’à ce jour, la plupart des anciens Zimbabwéens blancs ignorent que Ian Smith n’a jamais pris le risque de se présenter à une élection démocratique en tant que chef du parti. La plupart des gens n’avaient jamais entendu parler de Smith lorsqu’il avait assumé la direction d’un parti légitimement élu à la suite d’un coup d’État interne. Du jour au lendemain, Smith était Premier ministre, mais avant d’être reconnu par l’électorat en tant que chef du parti, il avait pris le contrôle des journaux, de la radio, de la télévision, de l’armée et du système judiciaire, se moquant de la démocratie et ridiculisant la plupart des Blancs. Les Zimbabwéens noirs et blancs opposés au racisme n’avaient d’autre choix que la guerre pour éliminer Smith et son parti. Ayant foulé aux pieds tout semblant de démocratie, Smith n’a jamais perdu un seul siège lors d’élections, le succès de son parti étant garanti par le contrôle des médias bien avant les élections. Le seul siège qu’il ne contrôlait pas dans un Parlement de 50 sièges était celui que j’avais détenu pendant un certain temps car, comprenant ce que Smith avait fait, j’ai secrètement pénétré dans son parti pour obtenir un siège, puis j’ai traversé le parquet pour former une opposition. Ma décision était si secrète que seuls Pat Bashford, chef du Parti du Centre multiracial détruit et Sir Roy Welensky, ancien chef du parti initial de Rhodésie, en étaient au courant à l’avance.
En résumé, je crois avoir donné des raisons suffisantes pour montrer que l’existence même du système de partis politiques hérité du Zimbabwe et d’autres pays africains exclut une bonne gouvernance. En outre, cela conduit à des violences inutiles lors des élections et ouvre nos nations à la corruption et à la possibilité d’une nouvelle et sinistre forme de colonialisme.
Les ouvriers du textile sont déjà inactifs et sans emploi dans les pays voisins, la Zambie et l’Afrique du Sud, et une pollution sans précédent s’ensuit alors que le corporatisme chinois s’installe, soutenu par le parti au pouvoir dans le gouvernement le plus corrompu jamais connu au Zimbabwe. Cela étant, permettez-moi de suggérer comment le Zimbabwe, la Zambie, l’Afrique du Sud ou tout autre pays pourraient former une démocratie sans parti plus honnêtement démocratique dans laquelle tous les citoyens pourraient se sentir plus libres, en sécurité et bien gouvernés.
Section III. Une démocratie sans parti et une bonne gouvernance
Mes observations et mon expérience m’ont convaincu que les partis politiques doivent céder la place à une démocratie sans parti si l’on veut que les gens connaissent un jour une bonne gouvernance. Tant que la démocratie sans parti ne sera pas instaurée, la bonne gouvernance restera une simple idée et ne sera jamais une réalité dont les gens pourront profiter.
Pour paraphraser Ghandi, la bonne gouvernance ne restera qu’une bonne idée jusqu’à ce que tous les partis politiques soient interdits.
III-A. Élire les députés.
Dans une démocratie sans parti, il serait évidemment nécessaire d’élire des députés parmi lesquels un gouvernement serait formé. Ci-dessous, je décris une manière dont le peuple pourrait élire ses représentants au Parlement. Ce n’est qu’une suggestion pour encourager la discussion. Ce n’est certainement pas le seul moyen, ni probablement le meilleur, d’organiser de telles élections.
Le pays serait divisé en circonscriptions électorales ou circonscriptions comme aujourd’hui, mais avec les limites des circonscriptions déterminées par le pouvoir judiciaire et non par les politiciens. L’objectif serait d’empêcher la manipulation des rôles des électeurs, des limites des circonscriptions électorales et bien plus encore par des groupes d’intérêts spéciaux dans le pays. N’importe quel individu pouvait se présenter aux élections si sa candidature était appuyée par un certain nombre de signatures de personnes résidant dans cette circonscription (supposons 2 000 à des fins d’illustration). Ces aspirants députés feraient appel au soutien des électeurs de leur circonscription sur la base de leur caractère, de leur réputation et de leurs antécédents dans la communauté et de leur désir de servir leur communauté et leur nation à ce titre.
Ils collecteraient leurs propres fonds auprès de leurs partisans, avec une limite stricte sur le montant que chaque individu pourrait contribuer, et se fieraient à leur mérite. Tout candidat au Parlement qui serait soutenu financièrement par une entreprise ou un groupe d’intérêt spécial serait disqualifié et ne serait plus jamais autorisé à se présenter à une charge publique.
Dans le cadre d’élections au cours desquelles les individus se présentent en fonction de leurs mérites plutôt que des dogmes des partis politiques sur des questions ou des politiques préfabriquées, il est difficile pour les forces armées de soutenir autre chose que l’État, comme elles ont juré de le faire constitutionnellement. Il y a un monde de différence entre un candidat qui se présente pour un parti, qui régurgite la politique du parti, qui bénéficie du soutien d’un chef de parti qui se porte garant de sa réputation, qui bénéficie d’un compte de dépenses couvert par des sociétés ou des fonds de son parti, et un candidat qui le finance ou le finance. elle-même tout en faisant appel à du soutien en fonction de ses capacités, de sa réputation et de sa position dans la communauté.
Les fonds nécessaires pour se faire élire par un individu dans sa circonscription sont infimes comparés aux fonds nécessaires aux partis politiques confrontés à des élections. Cela élimine à lui seul une source majeure de corruption, à savoir les groupes d’intérêts spéciaux qui financent les partis politiques pour influencer les élections. Faire en sorte que les citoyens soient élus selon leurs mérites dans leurs propres circonscriptions découragerait également la tendance à lutter contre les élections par le biais de la publicité télévisée, qui tend à privilégier l’apparence au détriment du fond. Il n’existe aucun lien connu entre la richesse et la sagesse, l’expérience, la personnalité équilibrée et l’intelligence requises pour se présenter à des fonctions politiques.
Un tel système électoral sans parti en Afrique supprime le chaos émotionnel qui résulte lorsque de grandes masses portant des T-shirts du parti, prononçant des slogans émouvants tout en brandissant des machettes et soutenues par la police et l’armée, exigent à tout prix la destruction de l’opposition et la victoire. Le besoin actuel de fraude électorale, d’achat de voix, de manipulation des limites des circonscriptions, de diffamation, d’intimidation, de meurtre et de torture disparaît avec l’absence d’un parti pour soutenir ou cautionner de telles actions. Et bien sûr, un tel comportement des partis politiques décourage gravement les investissements en capital, ce qui entraîne ultérieurement une perte financière d’indépendance et une vulnérabilité face au danger émergent du nouveau colonialisme d’entreprise.
Un autre avantage de ces élections sans parti est qu’elles éliminent les retards préjudiciables et la violence qui surviennent lorsque les partis politiques vaincus exigent un recomptage et un recours aux tribunaux. Si un particulier demandait un recomptage des votes dans sa circonscription, il n’y aurait pas de retard émotionnel dans la formation d’un gouvernement. Parce que le gouvernement n’est pas formé par un seul parti, il n’est pas nécessaire d’avoir une majorité au Parlement – la cause de tant de violence lors des élections.
Une telle élection d’individus de bonne réputation rend difficile aux groupes d’intérêts spéciaux ou aux entreprises de corrompre, de financer des partis ou de falsifier les élections de toute autre manière. Même si cela est vrai en principe, les superpuissances et les groupes d’intérêts spéciaux ne vont pas disparaître du jour au lendemain. Comme l’ont fait les compagnies de tabac, et comme continuent de le faire les sociétés pétrolières et charbonnières ainsi que les sociétés agrochimiques, elles persisteront dans leur recherche du profit à court terme tout en laissant la société supporter les véritables coûts environnementaux, économiques, sanitaires et sociaux. En raison du désir de puissantes sociétés transnationales de contrôler les ressources des pays sous-développés dans la nouvelle forme de colonialisme, nous pouvons nous attendre à la poursuite de campagnes massives de désinformation de la part de ces sociétés. Les entreprises avides de pouvoir, comme le léopard qui ne change pas de place, changeront simplement de tactique. Il est donc nécessaire d’avoir une constitution vivante pour répondre aux tactiques changeantes des intérêts particuliers.
III-B. Former un gouvernement
J’utilise des nombres arbitraires uniquement à des fins de facilité et d’illustration. Supposons qu’un État comme le Zimbabwe soit divisé en 200 circonscriptions basées sur la répartition de la population, dont les limites sont déterminées par le pouvoir judiciaire. Lors d’élections tenues tous les cinq ans, 200 députés seraient élus comme représentants du peuple. Ces députés formeraient alors un « collège électoral » pour élire un Premier ministre afin de former un gouvernement. Les membres aspirant à diriger et à former un gouvernement s’adresseraient à tous les députés et, après discussion, les membres voteraient pour la personne qu’ils jugeaient la plus capable et la plus apte à assumer ce poste. La personne ainsi élue comme Premier ministre procéderait ensuite à la nomination des personnes qu’elle choisirait comme ministres parmi les 199 députés restants. Chacun des choix de cabinet du Premier ministre est discuté et approuvé par un vote de 80 % de tous les députés et un gouvernement est ainsi formé. Il est suggéré d’exiger un pourcentage élevé de soutien des députés dans des cas tels que le Zimbabwe, car cela rendrait difficile à tout Premier ministre de former un cabinet tribal.
Les autres représentants du peuple au Parlement constituent des députés d’arrière-ban sur lesquels on peut faire appel pour les nombreuses commissions nécessaires et qui participeront naturellement aux débats et à l’approbation ou au rejet des politiques gouvernementales. Contrairement à la sélection des partis, cela sert à empêcher que les commissions parlementaires soient sélectionnées sur une base partisane prédéterminée, ce qui peut conduire à des conflits et conduire à la sélection de personnes pour des raisons autres que la compétence.
La nature humaine étant telle qu’elle est, nous pouvons toujours nous attendre à ce que des personnes tentent de manipuler le processus et de former des cliques de partisans à l’intérieur et à l’extérieur du Parlement, mais dans un système tel que celui décrit, cela ne prend pas les proportions ni les dangers inhérents au système des partis politiques. .
Des limites de mandats seraient appliquées à tout Premier ministre, les mandats étant prolongés uniquement avec une approbation très élevée spécifiée par la Constitution ; par exemple, 80 % des députés en exercice.
III-C. Destitution d’un gouvernement incompétent
Dans le cas où un gouvernement particulier se révélerait incompétent, tout ce qui serait nécessaire pour son destitution serait un simple vote de censure adopté par la majorité des membres du Parlement. Un gouvernement incompétent n’aurait aucune capacité à entreprendre les actions de contrôle et de manipulation que les gouvernements font couramment aujourd’hui pour rester au pouvoir à tout prix.
Si un gouvernement incompétent était renversé, il ne serait pas nécessaire de revenir à des élections nationales après un long retard. Les représentants du peuple au Parlement éliraient un nouveau Premier ministre parmi eux pour former un nouveau gouvernement, évitant ainsi les coûts élevés et les dommages à la nation qu’entraîne aujourd’hui un changement de gouvernement.
III-D. Destitution d’un député incompétent.
Avec le système des partis, de nombreux députés perçoivent un salaire élevé malgré leur non-performance et leur désintérêt évident pour les citoyens de leur circonscription (sauf lorsqu’ils font appel aux voix). Généralement, de nombreux députés suivent simplement le whip du parti et, lorsqu’ils ne sont pas tenus d’approuver la politique du parti, ils peuvent se rendre au barreau parlementaire. Dans une démocratie sans parti, tous les députés devraient être performants et prêter attention à leurs électeurs car ils pourraient être révoqués à tout moment par une pétition signée par un nombre important de personnes dans la circonscription (5 000 par exemple – nombre élevé pour qu’une telle action n’est pas frivole). Ce député sans valeur serait alors démis de ses fonctions et libre, s’il le souhaitait, d’affronter d’autres lors d’une nouvelle élection dans cette circonscription.
III-E. Empêcher que les questions soient résolues en secret sans débat public.
Une démocratie sans parti, telle que suggérée, élimine un autre mal du système des partis.
Il s’agit de décider de questions importantes dans l’obscurité des réunions secrètes du caucus du parti, avec la crainte de perdre la position et les privilèges du parti au premier plan dans beaucoup d’esprits. Ce n’est que lorsque des décisions ont été prises de cette manière, qui tue tout débat intelligent, que de telles questions sont soumises au Parlement pour un débat simulé suivi d’une approbation sans discussion par les députés du parti.
Ayant assisté à de nombreuses réunions secrètes hebdomadaires de ce type après avoir pénétré le parti au pouvoir comme je l’ai fait, je suis très conscient de combien un tel comportement de parti est antagoniste et contraire aux principes démocratiques. Rien ne peut remplacer le débat public pour préserver les intérêts des citoyens et garantir une bonne gouvernance.
Permettez-moi maintenant d’aborder d’autres ajustements structurels qui devraient être inscrits dans la constitution d’une démocratie sans parti pour remédier aux échecs actuels des États partis.
III-F. Un pouvoir judiciaire indépendant
La nécessité d’un système judiciaire indépendant est largement reconnue, même si elle est souvent violée, subtilement ou ouvertement, par de nombreux partis politiques au pouvoir. Quiconque doute de la capacité du système de partis à faire échouer les meilleurs plans n’a qu’à lire Gangs of America, de Ted Nace (Berrett-Koehler, 2005), dans lequel il décrit comment, malgré tous les efforts des pères fondateurs de l’Amérique pour assurer le Pour survivre à une république populaire, la volonté de grandes entreprises a finalement prévalu grâce au recours aux partis politiques et aux personnes nommées par la Cour suprême au fil du temps. Si les pères fondateurs avaient constitutionnellement interdit les partis politiques au lieu d’essayer de freiner leurs excès par l’intermédiaire de trois branches de gouvernement, peut-être que bon nombre des invasions américaines des pays voisins auraient pu être évitées.
Encore une fois, les États-Unis ne sont pas seuls, car la nature humaine est quelque peu prévisible. Les exemples de partis politiques arrivant au pouvoir et contrôlant peu après le système judiciaire abondent en Afrique. Au cours de ma vie, j’ai été témoin d’un tel comportement à deux reprises au Zimbabwe, sous deux partis totalement différents, l’un avant et l’autre après l’indépendance. Divers mécanismes pourraient garantir un système judiciaire indépendant, mais sans partis pour manipuler la sélection des représentants judiciaires, il y a plus de chances de maintenir cet idéal.
De même, en l’absence de parti capable de confondre loyauté entre parti et État, on pourrait plus facilement compter sur les forces armées pour défendre les citoyens en défendant la constitution et l’indépendance du pouvoir judiciaire et de la presse. Ce que beaucoup de gens ne réalisent pas, c’est la raison pour laquelle les Américains insistent sur le droit du peuple à porter les armes. En effet, tout au long de l’histoire, davantage de citoyens ont été tués par leurs propres armées, en raison de leur allégeance à des politiciens plutôt qu’à la nation, plutôt qu’à des ennemis étrangers. Les exemples historiques sont nombreux, notamment les armées chinoise et russe et le Zimbabwe en est un exemple moindre. Si nous regardons l’armée de ce qui est aujourd’hui le Zimbabwe, mais qui était officiellement la Rhodésie, nous constatons qu’elle a combattu pendant la guerre des Boers, la Première et la Seconde Guerre mondiale et en Malaisie, tuant peut-être quelques centaines ou milliers de soldats ennemis. Cependant, l’armée rhodésienne puis zimbabwéenne soutenant le parti politique racial de Smith, puis le parti politique de Mugabe, a tué environ 50 000 Zimbabwéens, dont 20 000 après l’indépendance.
Il n’y a guère de justification pour une grande armée permanente ou régulière au Zimbabwe, si ce n’est pour permettre au parti au pouvoir de réprimer les citoyens zimbabwéens. La nation ne peut pas se permettre une grande armée régulière incapable de la défendre contre un agresseur suffisamment puissant pour l’envahir au mépris de l’opinion internationale, et lorsqu’une petite armée régulière et une grande garde nationale ou une force citoyenne comme le Zimbabwe en a eu au cours des deux guerres mondiales sont tout ce qu’il faut. c’est nécessaire.
III-G. Faire face à la corruption dans une démocratie sans parti
La corruption prend de nombreuses formes, depuis les pots-de-vin mineurs versés aux fonctionnaires de bas niveau jusqu’aux pots-de-vin importants versés par les entreprises sous la forme de pots-de-vin aux fonctionnaires qui peuvent influencer la politique gouvernementale ou attribuer des contrats. La corruption, à un certain niveau, sera malheureusement présente pendant des années dans n’importe quelle société. Cependant, une corruption majeure de la part de groupes d’intérêts particuliers, d’entreprises et de gouvernements, qui n’est pas maîtrisée, constitue un danger pour des économies entières, peut conduire à des guerres et causer des souffrances massives qui vont à l’encontre de la bonne gouvernance. L’Argentine, autrefois classée parmi les pays les plus riches, a été réduite au statut de ce qu’on appelle le tiers monde en raison d’une effroyable corruption officielle.
L’économie du Zimbabwe continuera de décliner tant que le Zimbabwe restera l’un des pays les plus corrompus au monde. Les mesures qui suivent pourraient contribuer à garantir des contrôles stricts sur la corruption officielle la plus préjudiciable.
La constitution d’une démocratie sans parti prévoirait un bureau de médiateur permanent sous contrôle judiciaire. Un tel bureau permettrait de répondre légalement et à moindre coût aux préoccupations de tout citoyen victime d’un traitement injuste. Ce bureau serait également chargé d’enquêter et de poursuivre tous les cas de corruption possible, assortis de sanctions obligatoires sévères. Des sanctions sévères seraient imposées aux sociétés étrangères reconnues coupables de corruption, de lobbying ou de financement de candidats parlementaires. Ils pourraient, par exemple, se voir automatiquement retirer leur autorisation d’exercer leurs activités dans le pays et les responsables locaux impliqués purgeraient des peines de prison. Les entreprises locales ainsi reconnues coupables verraient automatiquement leur licence révoquée et leurs fonctionnaires emprisonnés jusqu’à ce que tout l’argent impliqué dans la corruption soit récupéré. Tout citoyen reconnu coupable de corruption serait automatiquement interdit à vie d’exercer une fonction publique et serait également emprisonné jusqu’à ce que tout l’argent impliqué soit récupéré, restitué à ses propriétaires légitimes ou confisqué au profit de l’État. Il n’est pas difficile de lutter contre la corruption si les partis politiques qui profitent de la corruption sont eux-mêmes rendus constitutionnellement illégaux dans une démocratie sans parti. Je crois que le système de partis perpétue une corruption majeure à laquelle un gouvernement sans parti n’aurait pas à faire face dans la même mesure.
Malheureusement, la plupart des pays africains qui s’orientent vers une démocratie sans parti entraîneraient avec eux un arriéré de corruption au sein des partis politiques et des entreprises, auquel le nouveau gouvernement devra remédier. La clé pour remédier à cet arriéré réside dans le fait qu’il faut toujours au moins deux personnes pour se livrer à un acte de corruption. Sachant cela, un moyen d’initier le changement est d’accorder un délai de grâce de six mois pendant lequel l’une ou l’autre partie à tout acte de corruption peut signaler au Médiateur et fournir des détails en toute confiance. Après la période de grâce, pendant laquelle personne ne saurait savoir qui a signalé un cas de corruption, des poursuites suivraient. Ceux qui dénoncent la corruption et fournissent des preuves ne pourront faire l’objet de poursuites, bien qu’ils soient tenus de restituer l’argent gagné grâce à leur implication. Si l’autre partie à la corruption ne déposait pas de plainte, elle s’exposerait à des sanctions complètes. De cette manière, de nombreux aveux seraient obtenus, ce qui faciliterait les enquêtes, les poursuites, la résorption des retards et le recouvrement des fonds volés. Dans tout pays où la corruption actuelle est à la fois officielle et endémique et où peu d’entreprises ou de familles peuvent rester à flot sans au moins des actes mineurs de corruption, l’exigence de déclaration initiale pendant le délai de grâce devrait être fixée à un niveau suffisamment élevé pour éviter de bloquer le système. avec de petits actes de corruption.
Ayant grandi dans un pays étonnamment exempt de criminalité et de corruption, puis l’ayant vu devenir l’un des pays officiellement corrompus au monde, je comprends à quel point cela est dévastateur pour la plupart des citoyens. Au Zimbabwe, la corruption et l’inflation ont siphonné des richesses obscènes au profit de quelques-uns, tout en détruisant les richesses, y compris les retraites, de la plupart de la population. J’ai également appris que le système des partis, combiné à l’incompétence de la fonction publique, obligeait pratiquement les entreprises et les entrepreneurs à se livrer à la corruption comme seul moyen de mener à bien leurs affaires. Supprimer le système des partis et assurer une bonne gouvernance, par le biais des mécanismes de fonction publique décrits dans la section IV, contribueraient grandement à éliminer une grande partie des raisons qui poussent les gens à s’engager dans la corruption. Une force de police professionnelle, correctement rémunérée et fidèle à la nation et à la constitution plutôt qu’à un parti politique contribuerait également grandement à lutter contre la corruption. Tant que le gouvernement de parti persistera, la volonté de prévenir la corruption officielle fera tout simplement défaut, car les partis politiques et les entreprises ont tous deux beaucoup à gagner à se soutenir mutuellement sous la table. C’est pourquoi, lorsqu’un nouveau parti est élu, la corruption persiste généralement.
III-H. Gouvernement local
Dans une démocratie sans parti, il faudrait veiller à ce que ce qui s’applique au niveau national s’applique également au niveau des collectivités locales. Dans de nombreux pays africains, cela s’appliquerait non seulement au gouvernement local des villes, mais également aux gouvernements ruraux traditionnels dirigés par des chefs. Actuellement, dans tous les pays africains, je suis conscient du fait que le système de partis, après l’indépendance, a continué la pratique coloniale consistant à priver les chefs de pouvoir en faveur du gouvernement central et du contrôle des partis politiques. Cette pratique, combinée aux politiques en matière de ressources naturelles mises en œuvre par les gouvernements et les agences de développement, contribue à détruire la culture et les moyens de subsistance des populations rurales, encourageant ainsi une nouvelle migration vers des bidonvilles surpeuplés.
Alors que certains chefs africains luttent pour maintenir l’ordre et préserver leur culture par l’intermédiaire de leurs tribunaux traditionnels, le système des partis et le degré de contrôle centralisé ont contribué à corrompre de nombreux chefs aussi gravement que les hommes politiques. Ainsi, les chefs ne devraient pas être à l’abri de poursuites intentées par le Bureau du Médiateur.
III-I. Poste de président.
De nombreux pays africains souhaitent inclure le poste de président, même si ce n’est que cérémonial, et il n’y a aucune raison pour qu’il ne soit pas maintenu dans une démocratie sans parti. Cependant, la personne occupant cette fonction serait soumise à toutes les lois applicables à tout citoyen. Un tel président pourrait être nommé conjointement par le Parlement et le pouvoir judiciaire à vie (ou jusqu’à sa retraite) et pourrait remplir de nombreuses fonctions utiles, ce qui en ferait un poste intéressant.
III-J. Honorer les partis politiques qui ont obtenu leur indépendance.
Après une longue guerre pour obtenir l’indépendance, comme dans le cas du Zimbabwe, de la Namibie ou de l’Afrique du Sud, il est compréhensible que les gens ressentent de la loyauté envers tout parti qui a joué un rôle majeur dans l’obtention de l’indépendance. Pour beaucoup, le fait qu’un tel parti perde le soutien du peuple en raison de son inévitable incapacité à assurer une bonne gouvernance et risque ensuite de perdre une élection témoigne d’un manque de loyauté envers le parti.
Heureusement, tout pays formant une démocratie sans parti, et mettant ainsi fin à tous les partis politiques, n’aura jamais à faire face à la possibilité d’une défaite aux élections du parti qui a obtenu son indépendance. La situation serait encore meilleure si le parti au pouvoir jouait lui-même un rôle majeur dans l’élaboration d’une constitution non partisane avec d’autres partis et organisations de la société civile. Dans une telle situation, le parti qui a accédé à l’indépendance serait historiquement honoré et ne serait jamais soumis à la défaite. Le parti ayant atteint son objectif aux yeux du peuple pourrait être honorablement retiré, un peu comme on retire un cuirassé qui a bien servi en guerre après que le besoin n’en soit plus présent. Je crois que le plus grand service rendu à ma nation que pourraient rendre ceux de ma génération qui ont été les dirigeants de notre longue et amère guerre pour l’indépendance serait de diriger un mouvement inaugurant une démocratie sans parti pour la véritable liberté, la démocratie et l’indépendance pour lesquelles tant de Zimbabwéens ont donné leur vie.
En supposant que nous ayons pleinement compris la nécessité d’une bonne gouvernance pour avoir un gouvernement capable de faire face à la complexité sociale, environnementale et économique dans l’élaboration des politiques. En supposant qu’une démocratie sans parti devait être formée dans le sens décrit dans cette section, pourrions-nous alors nous attendre à une bonne gouvernance ? Un pays dans lequel tous se sentent libres, en sécurité et bien gouvernés. La réponse dans Non.
Il existe un autre secteur du gouvernement qui exclut actuellement la bonne gouvernance : la fonction publique créée pour soutenir un gouvernement.
Section IV. La fonction publique
Peu après son arrivée au pouvoir, le président Mugabe a lancé un appel astucieux dans l’un de ses discours publics, dans lequel il a appelé les scientifiques à trouver des solutions aux nombreux problèmes auxquels son gouvernement était confronté car, comme il l’a dit, les politiciens ne pouvaient agir que sur les conseils des leurs conseillers scientifiques. Et quand les choses tournaient mal, c’étaient les politiciens et non leurs conseillers qui en prenaient la responsabilité. Ce que Mugabe n’a pas réalisé, c’est que si cela est vrai pour les scientifiques, cela l’est également pour la fonction publique.
La section I a présenté les nouvelles connaissances scientifiques qui expliquent pourquoi les scientifiques, moi y compris, ne pouvaient pas conseiller ou aider les décideurs politiques à faire face à la complexité impliquée dans toute politique traitant de la gestion des ressources. En raison du cadre de gestion réductionniste universel que nous utilisions, nous ne pouvions que conseiller le gouvernement sur la manière de traiter les nombreux symptômes de la désertification en Afrique – sécheresses croissantes, inondations, pauvreté, rupture sociale et violence – mais jamais sur la cause profonde de la désertification elle-même. .
Permettez-moi maintenant de revenir à la science des systèmes pour montrer ce qu’elle nous apprend sur le fonctionnement de toute fonction publique. La section 1 parlait de ce qui est défini comme des systèmes logiciels complexes (organisations humaines) étant complexes et auto-renouvelables. Des propriétés imprévues et inattendues apparaissent également, entraînant des problèmes extrêmement difficiles à résoudre. Des années d’expérience, étayées par la recherche, nous ont beaucoup appris sur les propriétés émergentes inattendues des bureaucraties de la fonction publique qui nous empêchent de parvenir à une bonne gouvernance.
La plupart des pays disposent aujourd’hui d’une fonction publique permanente qui gère leurs affaires quotidiennes. Dans certains pays, l’ensemble de la fonction publique reste intact après les élections. Dans d’autres, les hauts fonctionnaires de la fonction publique sont automatiquement remplacés par le nouveau parti, qui nomme les personnes en fonction de leur loyauté envers le parti, de leur soutien financier ou simplement de leur népotisme, plutôt que de leurs compétences.
Le président Mugabe, dès son arrivée au pouvoir, était servi par une fonction publique raisonnablement compétente, composée de Zimbabwéens noirs et blancs, et pendant un certain temps, tout s’est bien passé. La plupart de ces fonctionnaires avaient servi le gouvernement précédent avec suffisamment d’efficacité pour que le dollar zimbabwéen ait une valeur égale à celle du dollar américain après de nombreuses années de sanctions économiques mondiales. La mémoire des gens est courte, mais le Zimbabwe a été salué comme une réussite africaine grâce aux efforts du gouvernement de Mugabe dans le domaine de l’éducation et dans d’autres domaines. Son parti a ensuite pris la mesure que font de nombreux pays africains : « africaniser » la fonction publique. Cela signifie nommer des personnes à des postes de fonction publique sur la base de leur race, de leur tribu, de leur famille et de leur loyauté envers un parti. Au Zimbabwe, de bonnes personnes, généralement sans formation, sans expérience ou sans culture de service, ont occupé des postes en remplacement ou en remplacement de fonctionnaires expérimentés. Le manque de compétence dans la fonction publique a rapidement eu des répercussions sur l’ensemble de l’économie, entraînant une perte de confiance des investisseurs (il fallait parfois plus d’un an pour obtenir une réponse à une lettre commerciale) et de graves pertes d’emplois, suivies par l’inévitable montée en puissance d’un parti d’opposition. Ne croyant pas au concept d’une opposition loyale, toute la tragédie qui a suivi et que je n’ai pas besoin de souligner était presque inévitable.
Même si nous blâmons publiquement les individus à la tête de l’État, cette progression descendante était inévitable étant donné la baisse rapide de la qualité et de l’efficacité au sein de la fonction publique. Nous risquons de répéter ce cycle si les partis politiques continuent de récompenser leurs partisans en leur offrant des postes dans la fonction publique.
IV-A. Assurer l’efficience et l’efficacité de la fonction publique pour soutenir la bonne gouvernance.
L’idéal d’une fonction publique essentiellement permanente est né à juste titre et doit perdurer. Des bureaucraties telles que la fonction publique ont été développées à l’époque de Napoléon pour garantir leur efficacité. Ils visaient à améliorer l’efficacité en engageant des professionnels dûment qualifiés et instruits pour pourvoir les postes. L’intention était de mettre fin aux pratiques passées qui avaient conduit à des erreurs massives où les gens, plutôt que de gagner leur poste, en héritaient ou l’achetaient.
Ce sont les propriétés émergentes et imprévues des organisations humaines (systèmes logiciels), y compris des bureaucraties gouvernementales, qui constituent un problème sérieux pour toutes les nations. Même si ces bureaucraties parviennent effectivement à atteindre l’efficacité prévue, leurs propriétés émergentes imprévues conduisent généralement à la chute du gouvernement qu’elles sont censées servir, comme dans le cas du Zimbabwe.
Dans Les Bâtards de Voltaire, John Ralston Saul, qui a étudié les erreurs majeures à travers l’histoire, note que, quels que soient les qualifications, la brillance ou les bonnes intentions des individus d’une organisation, ils produiront souvent des résultats qui manquent de bon sens et d’humanité, et ils seront étanches. à de nouvelles connaissances en dehors de leur compréhension étroite.
Propriétés émergentes : manque de bon sens et d’humanité. Il s’ensuit que les politiques élaborées et mises en œuvre par la fonction publique manqueront également d’humanité et de bon sens, ce qui engendrera une mauvaise gouvernance. Inévitablement, c’est le parti au pouvoir, plutôt que la fonction publique, qui est
Les exemples de politiques de fonction publique manquant de bon sens et d’humanité sont nombreux. Par exemple, immédiatement après que des avions ont atterri sur les tours jumelles le 11 septembre, la Federal Aviation Administration des États-Unis a publié une règle interdisant aux pilotes privés américains de voler à moins de 10 milles d’une installation nucléaire. . Il s’agit d’une action raisonnable et compréhensible.
Ce qui manquait de bon sens, c’est que la FAA a refusé, malgré les demandes, de dire aux pilotes où se trouvaient ces installations, car il s’agissait d’informations secrètes impliquant la sécurité nationale. Ce manque de bon sens s’est encore aggravé lorsque l’association des pilotes a repéré tous les sites sur Internet. Encore une fois, la fonction publique américaine n’est pas seule. Des livres pourraient être écrits remplis d’exemples de règles et de réglementations provenant du Zimbabwe et d’autres pays qui manquent à la fois d’humanité et de bon sens. Même si je mets ici l’accent sur la fonction publique, la tendance des bonnes personnes employées dans les organisations à faire des choses stupides est bien plus large que n’importe quelle bureaucratie de la fonction publique. Demandez à n’importe quelle personne normale en Amérique s’il est logique que l’Amérique produise du pétrole pour cultiver du maïs afin de produire du carburant, et elle se moque d’une telle stupidité, mais l’Amérique fait exactement cela parce que des milliers de scientifiques travaillent dans des institutions.
Propriétés émergentes : étanches aux nouvelles connaissances. Lorsque les nouvelles connaissances entrent en conflit avec les croyances dominantes, les bureaucraties y résistent. Les individus au sein de n’importe quelle bureaucratie (université, organisme gouvernemental, organisation à but non lucratif) ne peuvent pas plus changer cette propriété émergente que n’importe quel étranger. Un cas souvent cité est celui de la Royal Navy qui a mis près de 200 ans, après la première démonstration, à accepter que le jus de citron préviendrait et même guérirait le scorbut, malgré le fait que la Grande-Bretagne perdait chaque année des milliers de marins à cause de la maladie. La marine marchande, également dirigée par des officiers brillants et dévoués, a mis encore 70 ans à accepter cette mission vitale.
Un exemple encore plus dévastateur est celui du surpâturage du bétail. Il y a environ soixante ans, un chercheur français, publié dans cinq langues principales, a établi que le surpâturage était fonction du moment et non du nombre d’animaux. Vitales pour inverser la désertification et pour la vie de milliers de personnes mourant dans des guerres désespérées et des génocides, ces nouvelles connaissances n’ont pas encore été institutionnellement acceptées soixante ans plus tard par aucune bureaucratie (gouvernement, université, ONG environnementale ou agence internationale). Des milliers d’individus au sein de ces bureaucraties ont accepté que le surpâturage n’est pas dû au nombre d’animaux, mais ils sont impuissants à changer la résistance aux nouvelles connaissances de leurs bureaucraties – qui ne change que lorsque l’opinion publique change de manière significative.
D’un autre côté, les bureaucraties, y compris la fonction publique, peuvent être parmi les premières à adopter une nouvelle pensée lorsqu’elle n’entre pas en conflit avec les croyances dominantes. Ils adoptent très rapidement les derniers ordinateurs ou gadgets électroniques. Rapide à utiliser la technologie pour construire des barrages qui ne pourront jamais lutter contre la désertification. Rapide à utiliser la technologie pour replanter des arbres qui ne peuvent pas inverser la désertification sur de vastes zones avec des précipitations inférieures à 500 mm. Tout ce en quoi la société croit ouvre la voie, mais inverser la désertification en utilisant la seule chose possible, à savoir un élevage correctement géré, a rencontré une forte opposition de la part des agences gouvernementales dans de nombreux pays, y compris le Zimbabwe.
Un autre problème récurrent des organisations est qu’elles admettent rarement avoir tort. Si une organisation fait quelque chose de mal, elle fera le tour des wagons et la défendra quel que soit le prix, et même cela va à l’encontre de sa mission même dans la vie. Le meilleur exemple est bien sûr la bureaucratie de l’Église catholique. Ils connaissent les prêtres pédophiles depuis des centaines d’années, mais plutôt que de défendre les innocents comme l’Église est censée le faire, ils ont protégé les prêtres. Les organisations ne se comportent tout simplement pas comme le ferait n’importe quel être humain normal.
Les propriétés émergentes imprévues sont presque impossibles à prévoir, celles évoquées ici n’ont été surmontées par la fonction publique d’aucun pays. Cependant, la bonne gouvernance exige que, même si nous ne soyons pas en mesure de résoudre de tels problèmes, nous devrions et puissions développer des structures pour les minimiser dans des organisations par ailleurs efficaces. Ci-dessous, je suggère des changements structurels qui, s’ils sont mis en œuvre, pourraient permettre d’atteindre cet objectif. Toutefois, avant de poursuivre les solutions, il reste quelques défis supplémentaires à relever au sein de la fonction publique.
IV-B. Processus de sélection négative dans une fonction publique permanente.
Un problème majeur est le processus de sélection négative à mesure que les gens montent dans le service. Je viens d’une famille de fonctionnaires et j’ai également servi au ministère des Colonies de Rhodésie du Nord, puis au Département du gibier de Rhodésie du Sud. À l’époque de mon père et à la mienne, les opinions divergentes de celles d’un supérieur n’étaient pas encouragées.
Des personnalités brillantes et très engagées ont certes accédé à la tête de départements, mais c’était plutôt l’exception que la règle. En général, ceux qui n’ont pas secoué le bateau se sont levés, tandis que ceux qui l’ont fait ont été forcés de partir ou sont partis frustrés. En travaillant avec un certain nombre de ministères et d’agences au cours des années suivantes en Amérique, en Australie, en Inde, au Canada, au Pakistan, en Namibie, au Lesotho, au Mexique, au Canada, en Afrique du Sud et dans d’autres pays, j’ai compris que ce que j’avais vécu au ministère des Colonies et au Zimbabwe est trop courant pour être ignoré si une fonction publique doit servir comme elle le devrait.
IV-C. Des lois donnant pouvoir à des réglementations faisant obstacle à la bonne gouvernance.
Lorsque les politiciens adoptent de nouvelles lois, il est courant de donner au ministre compétent le pouvoir de demander à la fonction publique d’élaborer les règlements qui guideront son application. Même si la loi est à juste titre débattue au Parlement, les règlements élaborés par la fonction publique font rarement l’objet d’un tel examen public. Avec une fonction publique, aussi brillante que soient les individus qui la composent, incapable d’éviter des résultats manquant de bon sens et d’humanité, il n’est pas surprenant que la pléthore de réglementations publiées aujourd’hui au Zimbabwe ait conduit à des souffrances humaines.
Une fois de plus, ce sont les politiciens qui portent la responsabilité de la mauvaise gouvernance qui en résulte, et non la fonction publique. Vivant comme je le fais également aux États-Unis, je suis pleinement conscient de la masse de réglementations souvent ridicules qui ont ainsi contribué à entretenir la frustration du public sur la base de laquelle le populiste Trump a accédé au pouvoir.
IV-D. Qui élabore la politique – les politiciens ou les fonctionnaires ?
Les hommes politiques vont et viennent tandis que la fonction publique assure la continuité. Théoriquement, les hommes politiques élaborent des politiques et la fonction publique les met en œuvre. En réalité, l’homme politique à la tête d’un portefeuille s’appuie sur les conseils des professionnels de ce ministère ou de cette agence. Même dans les cas où la contribution du public ou une expertise extérieure est sollicitée, tout cela est géré par la fonction publique. De cette manière, il est courant que le membre du Cabinet chargé, par exemple, de l’éducation, de la santé ou de l’agriculture, ait pour conseiller principal le chef de la fonction publique de ce département. Cette personne a généralement accédé à ce poste grâce à une sélection négative – sans offenser personne, sans faire de vagues, etc. Et cette personne dirige une bureaucratie de conseil et de mise en œuvre qui peut être en retard de cinquante à cent ans sur les nouvelles connaissances disponibles dans ce domaine. Les conséquences de toute politique élaborée par ce ministre sont inévitables, et la colère de l’électorat qui s’ensuit se déverse une fois de plus sur les politiciens et non sur la fonction publique. Dans les États multipartites dotés d’élections démocratiques, cette tendance se traduit par un changement constant de parti, convaincu que la prochaine fois ce sera mieux. C’est rarement le cas.
Il y a quelques années, j’ai organisé une formation de dix jours en gestion holistique pour les hauts fonctionnaires du ministère de l’Agriculture du Lesotho. Une fois qu’ils ont compris comment analyser la politique avec un cadre holistique, je les ai fait travailler sur la politique de conservation des sols du Lesotho. Ils ont conclu que leur politique augmenterait l’érosion des sols et mettrait ainsi en danger les rivières et les barrages de l’Afrique du Sud voisine. J’ai alors posé la question « Qui était responsable de l’élaboration de cette politique ? » Après avoir blâmé les politiciens, les médias et le public, une femme a finalement fait une déclaration révélatrice. Elle a dit au groupe : « Les politiciens vont et viennent. Regardez autour de vous. Nous avons tous les cadres supérieurs ici. Nous sommes le gouvernement. C’est nous qui élaborons ces politiques que les politiciens doivent approuver, et nous en sommes donc responsables.» En raison des propriétés émergentes imprévues mentionnées, le Lesotho continue de s’éroder à un niveau alarmant, mettant en danger les barrages sud-africains et bien d’autres encore.
Je crois que nous pourrions minimiser ces propriétés émergentes en structurant le gouvernement d’une manière différente de celle héritée de l’indépendance, et nous devons le faire, car le rôle de la fonction publique est crucial pour la bonne gouvernance.
Une façon d’y parvenir est d’ancrer dans la Constitution la structure nécessaire au maintien des rôles efficaces de la fonction publique, tout en surmontant les aspects que j’ai soulignés et qui présentent des problèmes. La structure que je suggérerais suit et est guidée par le contexte national holistique du Zimbabwe auquel j’ai fait référence plus tôt.
IV-E. Conseils consultatifs ministériels
Chaque portefeuille gouvernemental est dirigé par le ministre nommé par le Premier ministre. Chaque ministre, dès son entrée en fonction, formera un conseil consultatif ministériel. Le Conseil consultatif est présidé par le Ministre qui est tenu de nommer au moins 3 et au plus 5 (par exemple) conseillers extérieurs à la fonction publique à son conseil. Deux membres du Conseil doivent être sélectionnés pour leurs connaissances et leur expérience dans ce domaine et trois sans connaissances spécialisées dans ce domaine mais possédant une bonne formation en arts libéraux ou autre. Et des rendez-vous peuvent être pris depuis l’extérieur du pays si nécessaire. Comme mentionné précédemment, les meilleurs fabricants de bougies n’auraient jamais pu imaginer ou développer des lampes électriques, d’où la nécessité d’avoir des personnes ayant une large et bonne formation en arts libéraux dans tout conseil consultatif ministériel technique.
Sans plates-formes partisanes visant à biaiser la sélection de personnes, quelle qu’en soit la source, seul le mérite qualifie de tels conseillers ministériels. Le haut fonctionnaire du département en charge de ce portefeuille participe sur un pied d’égalité au conseil car il lui appartient de mettre en œuvre ultérieurement toute politique ou projet émanant du ministère. La fonction publique est ainsi représentée à titre consultatif mais ne peut pas bloquer la mise à disposition de nouvelles connaissances au ministre, ni dominer le ministre. Cette dernière situation est assez courante car le ministre est un profane et les fonctionnaires sont généralement des professionnels, bien que formés dans des fiefs étroits du savoir. Encore une fois, en examinant les recherches de John Ralston Saul sur l’augmentation des erreurs mondiales majeures suite au remplacement du leadership amateur par un leadership spécialisé hautement qualifié, il conclut : « La réalité est que la division du savoir en fiefs féodaux d’expertise a rendu la compréhension générale et une action coordonnée n’est pas simplement impossible mais méprisée et méfiante…. À bien des égards, les différences entre les différentes langues sont aujourd’hui moins profondes que les différences entre les dialectes professionnels au sein de chaque langue. Toute personne raisonnablement assidue peut apprendre une ou deux langues supplémentaires. Mais le dialecte du comptable, du médecin, du politologue, de l’économiste, de l’historien littéraire ou du bureaucrate n’est accessible qu’à ceux qui le deviennent ».
IV-F. Coordination des politiques et des projets.
Étant donné que la formulation des politiques et l’élaboration de projets constituent un rôle très important du gouvernement, toutes les politiques et tous les projets doivent être coordonnés de manière globale par le gouvernement. Qu’est-ce que cela signifie? La coordination holistique signifie que les politiques et les projets sont coordonnés sur les plans économique, social et environnemental. Une telle coordination n’existe aujourd’hui dans aucun pays. Actuellement, les gouvernements qui utilisent, comme tous le font, le cadre réductionniste universel pour formuler leurs politiques coordonnent leurs politiques et leurs projets à deux niveaux : la coordination politique conformément aux dogmes et aux convictions des partis, et la coordination économique à travers le processus budgétaire. La plupart, sinon la totalité, des débats portent sur le coût et l’objectif de toute politique ou projet, tandis que lors des séances du caucus du parti, la politique est alignée sur l’idéologie politique du parti.
Les exemples abondent de pays où des politiques et des projets atteignent leur objectif mais causent d’innombrables problèmes supplémentaires parce que la complexité impliquée n’a pas été prise en compte. La politique agricole américaine est un cas bien connu qui conduit involontairement à détruire les moyens de subsistance de milliers d’agriculteurs, non seulement aux États-Unis mais dans d’autres pays. La politique foncière du Zimbabwe visait à redistribuer les terres, contribuant grandement à l’effondrement de nombreuses entreprises urbaines et de l’économie nationale, ce que ceux qui ont conçu cette politique n’avaient clairement pas prévu.
Idéalement, tout gouvernement devrait non seulement vouloir connaître le coût, mais aussi s’assurer que toutes les politiques et tous les projets sont simultanément économiquement, socialement et écologiquement rationnels à court et à long terme, et donc susceptibles de faire face à la complexité d’une réalité sociale, environnementale et économique étroitement liée. Cela peut désormais être réalisé en alignant tous les projets ou politiques de développement sur un contexte national holistique. Une façon d’atteindre cette coordination nécessaire pourrait être de former un conseil de coordination comme suit.
IV-G. Conseil national de coordination des politiques holistiques.
Dans une démocratie sans parti, la coordination des politiques et des projets serait globalement plus complète grâce à un système constitutionnellement ancré. La coordination de toutes les politiques ou projets gouvernementaux potentiels relèverait de la responsabilité d’un ministre spécifique, soutenu par un Conseil de coordination des politiques holistiques mandaté par la Constitution, formé comme un Conseil consultatif ministériel mais doté d’une équipe de personnes formées à l’utilisation du cadre de gestion holistique. Ce ministère serait chargé d’analyser tout projet de politique ou de projet en utilisant le cadre holistique et le contexte holistique national. Aucun débat budgétaire n’aurait lieu sans un rapport à l’appui garantissant que les députés soient pleinement conscients des conséquences sociales, environnementales et économiques probables.
Élaborer une politique ou un projet judicieux
Parce que des politiques et des projets judicieux sont essentiels à la bonne gouvernance, j’aimerais donner un exemple de projet proposé pour le Zimbabwe et montrer comment il serait analysé de manière conventionnelle et holistique. S’il est aujourd’hui presque impossible d’élaborer des politiques ou des projets holistiques, cela devient relativement simple en utilisant le cadre holistique. Les responsables des gouvernements africains, indiens et américains ont pu le faire avec seulement dix jours de formation.
N’oubliez pas que la caractéristique commune des politiques et des projets est qu’ils ont toujours des objectifs et des actions déterminés par des experts pour résoudre un problème. Toujours sans exception, l’ensemble de la complexité sociale/culturelle, économique et environnementale est réduit au problème en tant que contexte/raison de la politique.
Comment la politique ou le projet serait formulé/développé de manière conventionnelle
Au Zimbabwe, la deuxième plus grande ville, Bulawayo, manque d’eau. Le gouvernement a pris conscience du problème et les fonctionnaires de la fonction publique ont été chargés d’y répondre par une proposition de politique ou de projet. Le projet qui a émergé a un objectif, qui est en l’occurrence d’approvisionner en eau la ville. Dans une situation difficile comme celle-ci, il est généralement courant que les gouvernements engagent des consultants (la Banque mondiale est généralement impliquée) pour travailler avec le ministère concerné. Entre consultants et fonctionnaires, la proposition est de construire un grand barrage de retenue dans un premier temps, pour un coût de plus de 500 millions de livres sterling. Par la suite, ils proposent que l’eau soit acheminée vers le barrage depuis le lointain fleuve Zambèze, ce qui nécessiterait une énergie et des dépenses considérables pour pomper l’eau. Le projet, comme toujours, atteindra clairement l’objectif en fournissant davantage d’eau. Le débat porte désormais principalement sur les aspects budgétaires et la capacité à emprunter les fonds nécessaires.
Les entrepreneurs et les fournisseurs tournent en rond comme des vautours autour d’une carcasse et les pots-de-vin affluent pour influencer les fonctionnaires et les politiciens du parti et le barrage est finalement construit, fournissant à la ville de l’eau supplémentaire. Objectif atteint.
Parce que nous sommes confrontés à une complexité, d’autres problèmes surgissent inévitablement – perturbations sociales, perte de terres agricoles, augmentation des maladies (bilharziose et paludisme), perturbation du débit des rivières, ensablement du barrage, augmentation de la migration urbaine et de la demande en eau, perte des paysages et des espèces par exemple et plus tard les problèmes associés à l’interférence du débit d’eau d’un fleuve international. Les politiques ultérieures seront sans cesse développées pour faire face aux problèmes qui se poseront, comme elles le feront sûrement selon un modèle désormais familier au monde.
En raison du pourcentage extraordinairement élevé de sol nu entre les plantes dans tout le Zimbabwe (désertification), le barrage finira par s’envaser et éclater, rendant la ville une fois de plus à court d’eau, ou toute l’eau devra être pompée du Zambèze, créant ainsi des problèmes internationaux. Ce schéma n’est ni simpliste ni exagéré, comme l’a montré l’analyse de centaines de politiques et de projets de gouvernements, de la Banque mondiale et d’autres agences internationales et ONG, et des livres entiers ont été écrits sur ces conséquences involontaires. Ainsi, les déserts continuent de progresser, l’aide internationale fait trop souvent plus de dégâts que de bien, la biodiversité continue de diminuer, les mauvaises herbes continuent d’envahir, les inondations et les sécheresses augmentent sans changement climatique, l’érosion des sols augmente tandis que la pauvreté, la violence et la déception des gouvernants perdurent.
Comment la politique ou le projet serait formulé/développé dans un État non-parti garantissant que le développement et les politiques sont conformes à un seul contexte national holistique.
Dans ce cas, lorsque le gouvernement prend conscience du problème de manque d’eau à Bulawayo, le ministre du Développement de l’eau (ou le nom actuel du portefeuille) charge son Conseil consultatif ministériel de donner des conseils sur les solutions possibles. Le Conseil consultatif ministériel, en utilisant le cadre holistique, détermine d’abord la cause du problème.
Aucun problème ne peut être résolu de manière permanente par une politique ou un projet sans s’attaquer à sa cause profonde. Diagnostiquer la cause du problème est toujours la première étape lors de l’utilisation du cadre holistique. Dans ce cas, le Conseil consultatif ministériel se rend compte que la ville est à court d’eau en raison de l’ampleur et du rythme de la désertification dans cette région du Zimbabwe. La désertification entraîne des pertes d’eau très élevées de deux manières : des crues soudaines massives dues à la mise à nu des sols (l’eau s’écoule simplement) ; et évaporation (jusqu’à 80 % sur sol nu, tel qu’exposé sur sol couvert.)
Les pertes annuelles de précipitations reçues dans cette région du Zimbabwe, selon la saison, peuvent dépasser 80 ou 90 %, ce qui représente des milliards de litres de plus que ce que la ville pourrait jamais utiliser. Que la ville manque d’eau n’est donc ni une surprise ni un problème : c’est simplement le symptôme d’une grave désertification.
À la suite de ce diagnostic, le Conseil consultatif ministériel se rend compte que la construction du barrage ne résoudra pas le problème. Non seulement cela ne résoudra pas le problème, mais cela entraînera d’autres problèmes à mesure que tous les barrages actuels se rempliront de limon. Le Conseil comprend que la construction des barrages serait une solution coûteuse à court terme, aggravant une situation déjà grave et mettant davantage en danger la ville tout en chargeant la nation de dettes inutiles. Ils se rendent également compte de cette analyse qu’il y aura d’autres coûts sociaux et environnementaux négatifs associés au barrage qui ne sont ni nécessaires ni exigés.
Fort de ces connaissances, le Conseil consultatif ministériel élaborerait une politique qui inverserait la désertification et améliorerait l’efficacité du cycle de l’eau dans toute la région du pays dans laquelle se trouve le bassin versant de la ville. Inverser la désertification et améliorer l’efficacité du cycle de l’eau sur cette vaste zone permettrait non seulement d’éviter l’envasement continu des barrages actuels, mais garantirait également le réapprovisionnement des aquifères souterrains et un débit plus permanent des rivières. Ensemble, cela équivaudrait à une nouvelle eau chaque année dépassant de nombreux barrages de la taille du barrage proposé sans qu’il soit nécessaire d’interférer avec le débit du fleuve international Zambèze. Le Conseil reconnaîtrait, comme peu de gens le font, que le plus grand réservoir d’eau douce sur terre est le sol – dépassant de loin tous les rivières, lacs et barrages du monde. En outre, le simple fait de s’attaquer à la désertification améliorerait le bien-être et la prospérité de tous les habitants de l’ensemble du bassin versant et de la ville, tout en apportant une solution permanente car le problème, plutôt qu’un symptôme, a été traité. Le coût de l’inversion de la désertification et de l’amélioration du cycle de l’eau ne représenterait qu’une petite fraction du coût du projet proposé, et il ne nécessiterait pas d’emprunts, de service de la dette ou de taxation supplémentaire de la population pour finalement payer cette folie.
À ce stade, le Conseil consultatif ministériel adopterait une politique visant à inverser la désertification. Tout ce dont le pays a besoin pour inverser la désertification est déjà disponible dans le pays. Les connaissances nécessaires pour y parvenir ont non seulement été développées dans le pays, mais sont disponibles depuis plus de cinquante ans, bien qu’elles aient été empêchées de devenir une politique à cause de la fonction publique adhérant à de vieilles croyances non soutenues par les progrès de la science. La politique visant à définir les actions requises de la part des services d’éducation et de vulgarisation actuels ne nécessiterait pas plus d’une semaine. Il s’agit d’un cas très simple impliquant la réduction de l’utilisation du feu et la formation des personnes à gérer un nombre croissant de bétail de manière à minimiser le surpâturage des plantes tout en couvrant le sol et en inversant ainsi la désertification. Au moment où j’écris, des chiffres sont arrivés sur une parcelle de terrain du Dakota du Nord suite à de telles pratiques où l’infiltration d’eau sur les sols supérieurs du bassin versant a augmenté de 775 % (de 20,32 mm par heure à 157,48 mm par heure de taux d’infiltration des précipitations).
Le cadre politique élaboré serait désormais transmis au Conseil de coordination holistique pour une évaluation indépendante et un rapport avant d’être transmis au gouvernement pour être présenté au Parlement pour débat avant sa mise en œuvre. Le Conseil consultatif ministériel et le Conseil de coordination holistique ont veillé à ce que la politique et toutes les actions qu’elle incarne soient conformes au contexte national holistique et donc simultanément économiquement, socialement et écologiquement rationnelles à court et à long terme. Le débat ultérieur se concentrerait sur les mérites, les coûts et les avantages de la politique, sans aucune influence des partis politiques ou des entreprises ayant soudoyé des fonctionnaires. Une telle politique serait au-dessus de la politique partisane et à long terme, comme tout devrait l’être, mais aujourd’hui peu le sont.
Bien que ce que je décris puisse sembler long, en réalité, analyser et élaborer des politiques ou des projets en utilisant le cadre holistique est plus rapide que la manière conventionnelle utilisant le cadre réductionniste universel. En Inde, les responsables forestiers, après une semaine de formation, ont pu analyser 12 de leurs politiques et projets actuels et prévus en moins de trente minutes et conclure que tous endommageraient les forêts indiennes et aggraveraient ainsi les problèmes sociaux, économiques et politiques. Rien n’a changé au sein du Service indien des forêts en raison des propriétés émergentes qui caractérisent ces bureaucraties.
IV-H. Politique foncière
Jusqu’à ce qu’une nation ait une politique foncière et agricole globalement saine, la bonne gouvernance à long terme est irréalisable en raison du lien entre la santé des terres et l’économie, la fréquence et la gravité des sécheresses et des inondations, la pauvreté, la stabilité sociale, la violence et le sort ultime. de la nation. Et au fait que sans agriculture (la production de nourriture et de fibres à partir des terres et des eaux du monde), il n’est pas possible d’avoir une église, une université, une armée, un parti politique, un musée, une ville ou tout autre élément de la civilisation urbaine. .
Le Zimbabwe a redistribué ses terres et d’autres pays africains voisins sont influencés négativement par cette action ou envisagent eux-mêmes des politiques similaires, comme le sont les politiciens sud-africains racistes. Il serait donc sage d’examiner la situation brièvement.
Au moment où le gouvernement du Zimbabwe a décidé de redistribuer les terres (en réponse au chômage massif et à la montée d’une opposition politique), tout le monde, y compris les agriculteurs commerciaux et le gouvernement britannique, soutenait cette nécessité. Comme pour le problème de l’eau de Bulawayo décrit, la politique de redistribution des terres du Zimbabwe était orientée vers un objectif : la redistribution des terres. Rares sont ceux qui ignorent que les résultats ont été catastrophiques sur les plans économique, social, politique et environnemental. Ce résultat était inévitable simplement parce que le gouvernement a réduit l’écheveau de la complexité sociale, culturelle, économique et environnementale au problème de la perte du soutien politique comme contexte de sa politique de répartition des terres. Si les agriculteurs commerciaux déplacés, les Britanniques ou tout autre gouvernement avaient adopté une politique de réduction similaire, le résultat, bien que moins immédiat et moins violent, aurait finalement été essentiellement le même.
En septembre 1996, j’ai écrit au président Mugabe pour lui demander d’aller de l’avant avec notre politique foncière, et lorsque finalement le gouvernement du Zimbabwe a annoncé qu’il allait de l’avant, un groupe de Zimbabwéens à Harare ont participé à des ateliers auxquels les fonctionnaires ont refusé d’assister. Dans ces ateliers, nous avons d’abord cherché à voir quelle sorte de politique foncière émergerait de la redistribution des terres de la manière habituelle pour déterminer diverses actions dans le contexte du problème. Il est vite devenu évident pour tous que cela conduirait à un désastre toujours plus grand. Ensuite, nous avons utilisé un contexte national holistique générique similaire à celui mentionné précédemment pour voir si le cadre d’une politique pouvait être développé. Ce qui en est ressorti a été éclairant dans la mesure où il nous a montré qu’une politique pouvait être élaborée avec des résultats différents, et comment la société civile devrait et pouvait être rapidement impliquée pour obtenir un large soutien national dans la mise en œuvre de cette politique. Une politique élaborée de cette manière aurait abouti à une augmentation considérable de l’emploi, à l’installation de deux ou trois millions de personnes sur la terre, sans perdre un seul agriculteur, à une augmentation de l’assiette fiscale du gouvernement et à l’inversion de la désertification des fermes et des ranchs (ce qui est sérieux même sur les meilleurs) ainsi qu’une augmentation de la production alimentaire.
Il est rarement vraiment trop tard. Même aujourd’hui, bien que de nombreux dégâts aient été causés en appauvrissant la plupart des Zimbabwéens, l’une des questions immédiates auxquelles tout gouvernement non parti devrait s’occuper serait à la fois la politique agricole et foncière. Et les gouvernements africains seraient généralement bien avisés d’envisager d’élaborer de telles politiques de manière holistique, car la violence induite par la dégradation des terres et l’augmentation de la population est en augmentation dans toute l’Afrique. Et partout en Afrique du Nord, jusqu’en Inde et jusqu’en Chine, cela entraîne beaucoup de violence et d’émigration, modifiant le visage politique de l’Europe.
IV-I. Des politiques qui nuisent aux nations africaines
Par souci de brièveté, je n’ai pas traité d’autres politiques, mais je mentionne que le Zimbabwe et d’autres pays ont des politiques actuelles impliquant des choses telles que l’interdiction de voter aux citoyens de l’extérieur du pays et l’interdiction de la double nationalité. De telles politiques insultent notre intelligence, même si nous devons les analyser. De toute évidence, le contexte d’une telle action politique était de maintenir le parti en place au pouvoir, sachant que la plupart des personnes forcées d’émigrer ne soutiennent pas le parti qui les a obligés à quitter leur pays. Bien que de telles politiques soient clairement conçues pour atteindre les objectifs politiques des partis au pouvoir, considérées de manière holistique avec un contexte national holistique comme guide, elles se révèlent vite manquer de bon sens et d’humanité et avoir des motivations sinistres. De telles politiques ne peuvent qu’être préjudiciables à l’investissement en capital et à la rétention des personnes qualifiées et donc préjudiciables à l’économie et à tous les Zimbabwéens. Chaque personne qualifiée retenue tend à créer des emplois et à développer une économie tandis qu’à l’inverse, la perte de personnes qualifiées entraîne des pertes d’emplois et une plus grande pauvreté pour tous.
IV-J. Évaluation de l’aide internationale
Les pays africains recevant l’aide de nombreuses agences de développement, groupes religieux et grandes organisations environnementales doivent garantir que cette aide n’interfère pas avec la bonne gouvernance. Après avoir analysé de nombreux projets d’aide, je reconnais que, bien que bien intentionnés et répondant souvent aux besoins humanitaires immédiats, le résultat global à long terme est décevant pour les donateurs et les bénéficiaires. Personne n’est responsable de cela, car les projets d’aide sont conçus pour résoudre des problèmes et ainsi toujours réduire la complexité du problème en question. La nature globalement mauvaise de l’aide n’est pas différente de la conclusion des professionnels américains selon laquelle la mauvaise gestion des ressources est universelle aux États-Unis, comme mentionné précédemment.
De plus en plus de gens commencent à se rendre compte que, malgré de nombreux projets et des millions de dollars, les choses ne s’améliorent pas. En fait, les faits suggèrent que l’aide fait actuellement plus de mal que de bien, ce qui n’a clairement jamais été son intention. Dès qu’un gouvernement sans parti dispose d’un Conseil de coordination holistique au niveau du Cabinet, l’une de ses fonctions devrait être d’évaluer toute l’aide étrangère afin de garantir qu’elle est holistiquement solide et conforme au contexte holistique national. Plutôt que de rejeter l’aide étrangère, cela entraînerait des modifications pour répondre aux intentions souhaitées des deux parties. Il est difficile d’exagérer l’impact de projets d’aide internationale et de développement défectueux. Les objectifs de développement durable (ODD) les plus récents des Nations Unies ont été salués dans le monde entier, mais une fois analysés, nous constatons que presque tous les 17 ODD s’attaquent aux symptômes de la désertification mondiale et qu’aucun d’entre eux ne s’attaque à la cause. Nous pouvons ainsi garantir une aggravation de la situation en Afrique, notamment en Afrique du Nord, en Inde et en Chine.
La surveillance de l’aide de la manière suggérée, combinée aux mesures proposées pour lutter contre la corruption officielle, contribuerait grandement à éviter que des millions de dollars d’aide occidentale ne se retrouvent dans des comptes privés suisses avec l’influence corruptrice des partis politiques permettant aux hommes en quête de pouvoir et de richesse facile. prospérer aux dépens des citoyens.
Conclusion
Je crois que la plupart des gens, y compris les hommes politiques de mon pays ayant de hautes motivations, aspirent à une vie telle qu’exprimée dans le contexte national holistique générique présenté plus haut et soutiendraient l’idéal d’une meilleure gouvernance grâce à une démocratie sans parti. Inévitablement, une minorité au pouvoir, ou aspirant au pouvoir et à la richesse facile grâce au système de partis, s’opposera constitutionnellement à la formation d’une démocratie sans parti.
Au moment où j’écris ces lignes, les partis politiques au Zimbabwe se disputent la position et le pouvoir par divers moyens, y compris le parti qui a refusé aux citoyens toute élection significative depuis l’indépendance et qui tente maintenant de changer de direction et de tactique pour restaurer le soutien.
Alors que les épées sont tirées et que de nombreuses blessures du passé sont encore palpables, un débat intelligent est difficile, voire inexistant, et au fil des années, nombreux sont ceux qui ont appelé à une intervention internationale.
Aucune solution imposée de l’extérieur ne durera et je crois fermement que nous, Zimbabwéens, de toutes races, tribus, sexes, cultures et croyances, devrions façonner notre propre salut. J’espère que ce que j’ai écrit et suggéré ici suscitera un débat plus ouvert et n’offensera aucun individu ou parti, ce qui n’était pas mon intention.
À la suite d’un débat à l’échelle nationale, s’il semblait logique pour la plupart des gens que la démocratie ne peut réellement fonctionner que lorsqu’elle est débarrassée du mal des partis politiques, alors le changement pourrait, je crois, être opéré rapidement et pacifiquement. Cela pourrait se faire simplement en ignorant les partis lors des élections et en élisant des députés indépendants favorables à l’interdiction des partis politiques. Dès que ces indépendants sont majoritaires ou qu’un parti ne peut former un gouvernement sans eux, ils sont effectivement au pouvoir. À ce stade où les indépendants détiennent la majorité, ils peuvent modifier la constitution comme l’ont fait les partis, mais ils interdisent désormais tous les partis politiques et organisent de nouvelles élections dans une démocratie sans parti.
En tant que personne apolitique, mais Zimbabwéen passionnément patriotique et loyal, je ne peux que souhaiter à mon peuple du succès dans les années à venir, ce que je ne vivrai pas assez longtemps. Et j’espère sincèrement que les discussions émanant de ces idées aideront d’autres nations, y compris nos voisins proches, à rechercher une bonne gouvernance.
Institut Salé
Le Savory Institute a pour mission de régénérer les prairies du monde et les moyens de subsistance de leurs habitants, grâce à
une gestion holistique . Depuis 2009, le Savory Institute dirige le mouvement de l’agriculture régénérative en équipant les agriculteurs, les éleveurs et les communautés pastorales pour régénérer les terres dans des contextes culturellement pertinents et écologiquement appropriés.
Le Manque d’Expertise Terrain en Agronomie : Un Défi Croissant et les Solutions de SCV Agrologie
L’agriculture fait face à des défis majeurs liés à un retard agronomique et un manque d’expertise terrain. Ce déficit entraîne des sols appauvris, une dépendance accrue aux intrants chimiques et une faible résilience face aux changements climatiques. Pour remédier à ces enjeux, des approches innovantes comme celles développées par SCV Agrologie offrent des solutions durables et adaptées aux réalités locales.
Les Conséquences du Manque d’Expertise Terrain
Dégradation des sols : l’Approche et mode de gestion des sols actuels favorise une perte de matière organique (bilan carbone négatif), l’érosion et la compaction des sols, réduisant leur fertilité à long terme.
Dépendance aux intrants chimiques : L’absence d’une approche agroécologique préventive pousse à une utilisation excessive d’engrais et pesticides, augmentant les coûts et l’impact environnemental. Faible adaptation aux conditions climatiques : Le manque d’innovation et de recherche-action limite la capacité des agriculteurs à adapter leurs pratiques aux aléas climatiques croissants.
Baisse de la rentabilité des exploitations : Sans accompagnement technique de qualité, les producteurs subissent des coûts élevés et une rentabilité de plus en plus précaire.
SCV Agrologie : Une Réponse Concrète à Ces Défis
Depuis 15 ans, SCV Agrologie accompagne les agriculteurs dans l’adoption des Systèmes de Semis Direct sur Couverture Végétale Permanente (SCV), une approche qui améliore la résilience des sols et la productivité agricole. Implantation des mélanges multi-espèces : SCV Agrologie a mis en place des mélanges de 12 à 20 espèces adaptées aux conditions pédoclimatiques québécoises, permettant une meilleure couverture végétale et un enrichissement du sol.
Expérimentations en conditions réelles : Contrairement aux recommandations génériques, SCV Agrologie mise sur une approche recherche-action-diffusion, testant et ajustant les pratiques directement avec les agriculteurs. Formation et accompagnement technique : Par des rencontres terrain et des échanges avec un réseau d’agriculteurs pionniers, SCV Agrologie assure un transfert efficace des connaissances et un suivi personnalisé. Optimisation des systèmes de production : L’intégration des SCV en cultures de pommes de terre et en systèmes fourragers a démontré des bénéfices en termes de réduction des coûts et d’amélioration de la santé des sols.
Un Modèle d’Agriculture basé sur le stockage du carbone Pour l’Avenir
Le retard agronomique peut être rattrapé grâce à des initiatives comme celles de SCV Agrologie qui misent sur l’innovation, l’adaptation locale et la diffusion des savoirs. Pour garantir un avenir agricole durable, il est crucial que les acteurs du secteur reconnaissent l’importance de l’expertise terrain et adoptent des approches éprouvées comme le SCV.
Une méthode pour réduire l’usage des pesticides gagne du terrain dans l’Est
Accéder à la section commentaires
« L’objectif de la cohorte, c’est que les producteurs puissent adopter l’un des principes du système de semis directs sur couverture végétale permanente. Parmi les principes, c’est l’absence de travail de sol, la biodiversité et la couverture végétale », explique l’agronome Louis Pérusse.
PHOTO : RADIO-CANADA / VÉRONIQUE DUVAL
Publié le 26 juillet 2023 à 1 h 46 UTC+2
Des producteurs agricoles du Bas-Saint-Laurent et de la Gaspésie implantent une méthode de gestion durable des sols sur leurs terres agricoles depuis un an : le système de semis directs sur couverture végétale permanente (SCV).
L’agronome Louis Pérusse implante le système de semis directs sur couverture végétale permanente, aussi nommé la méthode SCV, depuis une quinzaine d’années au Québec.
J’ai amené des producteurs à réduire le travail de sol pour certains à 100 %. On a des réductions de pesticides jusqu’à 70 %. Des engrais, la même chose Une citation de Louis Pérusse, agronome
L’approche mise sur l’augmentation de la qualité des sols et du rendement des cultures en faisant une meilleure rotation et en privilégiant la diversité des plantes, ce qui réduit l’usage des engrais et pesticides.
On veut vraiment créer un écosystème à la ferme basé sur le concept du génie végétal, c’est-à-dire utiliser les fonctions des plantes qui vont rendre les nutriments du sol plus disponibles aux cultures de récoltes. C’est recopier l’image du système forestier, mais au niveau agricole, explique l’agronome.
Ce n’est pas une approche qui exclut les engrais de synthèse ou les pesticides, mais l’objectif c’est de les réduire le plus possible, ajoute-t-il.Ouvrir en mode plein écran
Leurs fermes bovines ou laitières sont situées entre Rivière-du-Loup, dans le Bas-Saint-Laurent, et Caplan, en Gaspésie, avec une majorité dans la vallée de la Matapédia.
PHOTO : RADIO-CANADA / VÉRONIQUE DUVAL
La méthode SCV a été mise au point en milieu tropical il y a 40 ans. Elle a gagné en popularité ailleurs dans le monde au cours des 20 dernières années.
Faire des associations, garder des sols verts en permanence, ce n’est pas une pratique que j’ai inventée. C’est une pratique que j’ai initiée. Puis j’ai eu la chance d’avoir un mentor, Lucien Séguy, qui a travaillé un peu partout sur la planète, qui m’a transmis son savoir-faire, explique Louis Pérusse.
Son objectif depuis une quinzaine d’années est de former des producteurs dans cette approche globale d’agriculture durable.Ouvrir en mode plein écran
« On parle d’agir sur des enjeux comme l’érosion de sol, protéger le sol, nourrir le sol différemment que juste par des engrais de synthèse », explique l’agronome Louis Pérusse.
PHOTO : RADIO-CANADA / VÉRONIQUE DUVAL
L’idée de la méthode, c’est de ne plus labourer le sol.
Dans le fond, l’image, c’est que je veux avoir des champs verts le plus possible toute l’année. Je veux qu’au printemps ce soit vert. Je veux que ça soit vert à l’automne. Donc le SCV, c’est un peu l’image d’avoir une prairie en permanence, mais avec des cultures de récolte, explique-t-il.
Des agriculteurs préoccupés par l’avenir
Agriculteur pendant 45 ans dans la vallée de la Matapédia, Gilles St-Laurent est à présent à la retraite, mais il est toujours passionné d’agriculture.
Il accompagne la cohorte d’agriculteurs de la vallée de la Matapédia, de Rivière-du-Loup, de Caplan et de Cap-Chat qui se sont engagés à intégrer ces nouvelles pratiques.
Les agriculteurs sont très préoccupés actuellement par l’environnement et ils cherchent des solutions. Quand on voit les feux de forêt, quand on voit les pluies torrentielles qu’on a, il serait temps qu’on se réveille, déclare-t-il.Ouvrir en mode plein écran
« Il y a des changements climatiques. Qu’est-ce qu’on fait? Moi, je ne compte pas sur le gouvernement pour faire des changements, je pense que c’est dans chacune de nos entreprises, individuellement, qu’on peut faire des choses », affirme Gilles St-Laurent.
PHOTO : RADIO-CANADA / VÉRONIQUE DUVAL
Il explique qu’au cours des huit dernières années, même avec l’ajout de pesticides, d’engrais chimiques et d’herbicides, les rendements n’étaient plus au rendez-vous.
On se disait : il y a quelque chose qui se passe, il y a quelque chose qui se passe dans la terre, qui fait que même si on met plus d’engrais, ça n’augmente pas la production, poursuit M. St-Laurent.
Il estime que la technique SCV ne règle pas tous les problèmes en agriculture, mais qu’il s’agit d’une solution applicable dans la vallée de la Matapédia.
Gilles St-Laurent souligne que les producteurs agricoles utilisant la méthode SCV sont passés de 4 à 17 entre 2021 et 2023.
Il estime à près de 1200 hectares les terres utilisant le SCV dans la vallée de la Matapédia depuis 2021.
Seul, on va loin, mais en équipe on va plus vite, affirme l’agronome Louis Pérusse.
Il explique que le but de former des cohortes est de faciliter le transfert des connaissances et d’améliorer le réseautage entre les producteurs qui veulent implanter le SCV dans leurs pratiques.
Les participants à cette cohorte reçoivent un soutien financier provenant du Plan d’agriculture durable du ministère de l’Agriculture, des Pêcheries et de l’Alimentation (MAPAQ).
Élever notre sol maintenant, le nourrir pour se nourrir demain.
Notre ferme céréalière est en SCV depuis 25 années, je partage cette aventure avec mon épouse Lydie et maintenant ma fille Lucie lesquelles partagent avec moi la passion du SCV.
Cette ferme se situe dans le centre de la France près de la ville de Nevers. On y pratique donc l’Agriculture de Conservation des Sols (ACS)* (aucun travail mécanique du sol + rotations de cultures importantes + couverture permanente du sol avec résidus et plantes de services ) + apports de matières organiques exogènes .
Nous avons eu la chance de rencontrer M. Lucien SÉGUY , chercheur au Cirad, qui a travailler principalement au Brésil sur le SCV, mais aussi un peu partout dans le monde…..Avec Lucien, nous avons mis en place sur notre ferme, une plateforme d’essai spécifique consacrée principalement aux plantes de service .
Le reste de la ferme est donc dédiée aux SCV de M. Lucien Séguy, Notre objectif est aussi d’ élever le sol qui nous est confié….
M. Lucien Séguy :
Docteur en Agronomie, Scientifique du CIRAD, pédologue de l’ORSTOM, Lucien estimait que tous les outils sont connus pour produire intensivement, à peu de frais, de la nourriture de qualité sur des sols à fertilité améliorée. Lucien Séguy, est certainement l’Agronome qui, sous tous les climats,toute les latitudes, a sillonné, foulé de ses propres pieds, le plus de parcelles agricoles au monde et ce record n’est certainement pas prêt d’être battu !
Le CIRAD (*équipe Lucien.Séguy, ingénieur agronome, pédologue IRD, consultant international pour les systèmes de culture SCV, S. Bouzinac, S.Boulakia, F. Tivet, Hoa Tranquoc, R. Michellon, F. Jullien) et ses collaborateurs du Sud Asie ont créé au cours des 30 dernières années de nombreux scénarii diversifiés de développement durable en SCV de plus en plus performants.
L. Séguy et S. Bouzinac les ont créés/maîtrisés avec leurs partenaires au Brésil puis transférés/adaptés en France depuis le début des années 2000. Ces systèmes SCV, que l’on pratique en Bourgogne depuis 25 ans , sont issus de l’ingénierie écologique au service du développement et qui fonctionnent à l’image de l’écosystème forestier dont ils sont inspirés (biomimétisme), ils ont été continuellement perfectionnés au cours des 40 dernières années aux plans écologiques, agronomiques et technico-économiques.
Ils offrent, aujourd’hui, toutes les garanties de l’agriculture durable : De plus en plus productifs avec de moins en moins d’intrants chimiques, donc des coûts de production en baisse. Ils sont tous construits sur une reconquête de la biodiversité fonctionnelle : Rotations de cultures, intégration agriculture – élevage, sols toujours protégés sous couvertures mortes et/ou vivantes ; biologiquement très actifs, ils séquestrent efficacement le carbone :
Dans la plupart des fermes françaises qui les pratiquent depuis plus de 20 ans, le taux de séquestration annuel de carbone est compris entre 1 et 1,5 T/ha/an, soit 10 fois supérieur à la démarche « 4 pour 1 000 » ; de même, ils favorisent la rétention des nutriments (CEC plus élevée), réduisent l’incidence des maladies, des ravageurs en général, des nématodes phytophages, fonctionnent en circuit fermé comme la forêt et réduisent ainsi très fortement les pertes de nutriments (recyclage profond des bases et nitrates, injection de carbone en profondeur, hors des atteintes anthropiques) et garantissent la qualité biologique des sols, des eaux et des productions (système auto-épurateur , forte capacité de biorémédiation).
En forêt, rien ne se perd !
Tout comme Louis Pasteur, Lucien Séguy a passé une partie de sa jeunesse à peindre, exerçant et développant ses talents d’observateur. En grand scientifique, il interroge sans cesse la nature : « Mon maître, mon juge, mon centre d’inspiration principal, c’est la nature, sous sa complexité la plus grande. Vous avez une question à poser ? Posez-la à la nature. Et elle vous répondra ! Il faut simplement faire quelques manip’ pour qu’elle vous réponde. Et le faire de manière scientifique pour comprendre pourquoi cette réponse ; ce qu’elle veut dire exactement. »
Le but de Séguy est d’inventer une agriculture inspirée de la fertilité naturelle des forêts. Il fonde les principes de cette nouvelle agriculture sur quelques observations.
Les SCV (Semis sous Couverture Végétale) Vers du vert 365 jours / 365
Solution d’avenir pour une agriculture à très hautes performances économiques et environnementales …..et qui produit.
Les SCV, le génie végétal….. Qu’est que c’est…Qu’apporte t-il ?
Les SCV ouvrent maintenant la voie (et sont actuellement très proches) d’une gestion totalement biologique des agrosystèmes dans un environnement parfaitement protégé, avec ou sans apport de matière organique exogène. Nous savons aussi aujourd’hui, dans cette agriculture écologiquement intensive, comment supprimer le glyphosate et les pesticides, les intrants chimiques en général tout en maintenant de très hautes productivités plus stables dans le changement climatique. Cette reconquête de la biodiversité ramène l’évolution des systèmes cultivés vers celle des écosystèmes naturels originels (Résilience).
Ces dernières années, sur le réseau « SCV France informel » qui fonctionne sans aucun appui financier ou subvention, a été précisée/ajustée l’utilisation optimale du « génie végétal » au profit des performances des cultures, des sols et de la qualité biologique des productions : Les sojas conduits au Brésil avec ces techniques contiennent 3% de plus de protéines que les sojas conventionnels en semis direct. Le « génie végétal » est le pilier des systèmes SCV, et SCV BIO en construction .
Le SCV est matérialisé par des MIX de plantes de couverture installés ou entre les cultures ou en association avec elles, (en semis direct ou en semis à la volée qui réduit drastiquement l’incidence des adventices) dont la composition pluri-espèces est optimisée pour bénéficier d’un maximum de fonctions agronomiques gratuites au profit du système sol-type de culture ….
Ce génie végétal assure 90 à 95% des performances des systèmes : Productivités optimisées élevées et stables, contrôle naturel des adventices (système de double couverts successifs tournant dans la rotation , densité de semis élevée) recyclage très important de nutriments le + souvent supérieur aux besoins des cultures (excepté N sur céréales à compléter par faible doses de N organique) …. Perte minimum à nulle de nutriments dans le système sol-culture …forte capacité de biorémédiation (système épurateur des sols) …forte séquestration de Carbone (le sol devient maintenant un puits de carbone important…. Il devrait devenir un choix stratégique des gouvernements dans la lutte contre le réchauffement climatique … Le prix des terres devrait être fixé sur leur teneur en matière organique pour inciter les agriculteurs à pratiquer les SCV et SCV bio) … L’agriculture n’est plus un système de prédation mais devient un système de régénération de la fertilité globale des sols (optimisation de nombreuses fonctions agronomiques gratuites, services écosystémiques qui substituent la chimie intensive exogène et coûteuse)…
C’est la première fois dans l’histoire de l’agriculture que l’homme peut produire beaucoup, à peu de frais, tout en augmentant la fertilité organo-biologique de son sol …Ce mode de gestion écologiquement intensif permet ainsi de substituer progressivement l’utilisation massive actuelle d’énergie culturale d’origine industrielle par une énergie culturale d’origine biologique de plus en plus performante. Le patrimoine sol est cousu par les systèmes racinaires, l’érosion totalement contrôlée; nos sols, eaux, rivières et productions, sont « propres », nos paysages préservés.
La température est régulée comme sous forêt même au cours de canicules (régulation de l’ évaporation), infiltration maximum de l’eau de pluie (stockage) au détriment du ruissellement , d’où contrôle externalités, arrêt des coulées de terres , protection des infrastructures dont fossés de drainage, routes ….produits phyto, nitrates, non entraînés rapidement vers cours d’eau , la mer…
Cette approche, est obligatoirement intéressante pour les assureurs qui seraient très concernés par l’économie des catastrophes climatiques auxquelles ces compagnies sont maintenant confrontées. On pourrait même leur demander de participer financièrement à la diffusion de ces techniques plutôt que de payer des sommes astronomiques pour « tenter de réparer les dégâts »…qui se multiplient ….imprévisibles et croissants…
Cette nécessité de production totalement propre et de qualité constitue un élément de conviction puissant et démultiplicateur pour l’adoption/diffusion des systèmes SCV …C’est par l’adhésion, l’appui effectif des autorités (adhésion politique, promotion officielle , subventions) et de la société civile que l’appropriation des SCV pourra progresser le plus rapidement chez les agriculteurs .
Les SCV permettent de répondre à la demande sociétale actuelle d’amélioration de la qualité de l’alimentation et les SCV sont une solution à la problématique « eau » (érosion, ruissellement, qualité, efficience).. + Forte séquestration du C dans les sols (label bas carbone)
Et pour demain….En SCV, les sols accumulent les performances au fil des années, on peut donc encore améliorer fortement nos performances en SCV ….!
M.SÉGUY , nous ayant quittés le 25 Avril 2020 , nous tenons absolument à prolonger son œuvre , pour cela, un webinaire international a été organisé le 24 et 25 janvier 2023 :
Nous avons constituer, avec un groupe d’agriculteurs en SCV, une association à but non lucratif dédiée à l’agriculture de conservation des sols et plus particulièrement au Semis sous plantes de Couvertures Vivantes (SCV)
Cette association (SCV LUCIEN SEGUY) a pour but de former les nouvelles générations d’agriculteurs à ces technologies de production agricole innovantes, de tester de nouvelles techniques d’associations de plantes de couverture, d’expérimenter de nouveaux matériels en cours d’élaboration sur notre ferme et qui concernent la problématique « glyphosate » de ces techniques ACS, nous pensons pouvoir déboucher rapidement sur une solution intéressante et innovante.…
L’objectif de cette association, étant aussi de fédérer les organismes français et européens existants dans cette approche pour une meilleure efficacité nationale…voir internationale ….
On pense qu’il y a une certaine urgence à développer ces méthodes innovantes : La qualité, la durabilité des sols de notre planète deviennent préoccupantes pour la sécurité alimentaire et notre climat…, les coûts afférents au maintien de notre production conventionnelle actuelle commencent à présenter de vives inquiétudes de la part de nos collègues agriculteurs.
Par contre, nos systèmes de cultures SCV sont plus productifs, plus stables, plus attractifs économiquement et de moindre risque. Ce sont aussi ceux qui séquestrent le plus de carbone. Dans ces systèmes, la part de la fertilité gratuite construite en Semis Direct par voies physiques et organo-biologiques prend de plus en plus d’importance au cours du temps dans la capacité de production du sol : La productivité augmente avec moins d’intrants chimiques (engrais, pesticides), le potentiel du sol s’accroît, les coûts de production baissent et les impacts de l’activité agricole sur l’environnement sont mieux contrôlés.
Le semis direct sur couverture permanente du sol (SCV) est probablement le paradigme le plus complet qui ait été construit à ce jour pour le développement planétaire d’une agriculture durable, préservatrice de l’environnement, gérée, de plus en plus, ”au plus près de l’écologie”.
Nous sommes à un tournant historique dans la vie de l’humanité, les coûts environnementaux du développement économique dus aux énergies fossiles faciles, ont été largement ignorés , il serait dangereux de continuer dans cette voie.
L’humanité est maintenant exposée à un risque extrême du fait de l’incapacité de l’économie à prendre en compte l’épuisement rapide du capital naturel et doit trouver de nouvelles mesures de succès pour éviter une catastrophe.
La planète est notre maison, une bonne économie exige que sa gestion soit complètement revue ou même révolutionnée.
Ce constat sur l’ampleur des changements climatiques implique de revoir en profondeur notre agriculture, mais pas que ….!!
Plusieurs thèmes incontournables : En préalable : la globalité
A ce niveau de la présentation, il est évident que je vais vous évoquer le sol, la priorité des sols vivants, mais en même on est obligé d’aborder l’aspect globale des choses et les interactions qui les constituent ….Chaque élément est important mais tous ont des rapports incontournables entre eux et seul, un élément n’est pas fonctionnel.
L’énergie.
Le reste de nos énergies fossiles encore disponible doit pour une part contribuer à mettre en place de nouvelles solutions sobres et efficaces pour nos économies, ainsi que d’autres techniques de production énergétique sans gaspillage.
La technique SCV, par exemple, permet une économie de carburant fossile de plus de moitié permis pas le Non Travail du sol mécanique, en comparaison avec les techniques conventionnelles
On se libère du travail mécanique du sol, car celui-ci retrouve une résilience avec les systèmes racinaires permanents ….En SCV , la structure du sol est continuellement performante grâce à la vie biologique, le travail mécanique du sol n’ a plus sa place, les structures grumeleuses sont maintenues en permanence……et c’est bien ce travail mécanique qui est le plus coûteux en matière d’énergie, ce travail mécanique mis en place historiquement par nos anciennes générations d’agriculteurs pour gérer les plantes adventices n’a pu lieu d’exister en SCV…. !!
L’énergie d’hier, ça a été le soleil
L’énergie d’aujourd’hui,c’est le soleil…
L’ÉNERGIE DE DEMAIN, CE SERA LE SOLEIL ….Il y a une usine qui est super performante pour capter cette énergie encore gratuite…c’est la végétation et son procédé breveté c’est la photosynthèse..…la Nature phénoménale est même capable de stocker cette énergie solaire dans la puissance du génie végétal et le carbone ….Les graines et autres plantes que nous consommons sont bien des stocks d’énergie provenant des rayons solaires …..
Nous aurons le plaisir de vous accueillir sur Terres INNOVATION, la plateforme qui mettra en avant les innovations et des solutions agricoles pour accompagner toutes les Agricultures (grandes cultures, élevage, arboriculture, légumes, viticulture etc.) dans la transition agroécologique, la préservation des Biens communs au service de la performance des exploitations agricoles et de la pérennité des filières présentes sur le territoire.
Le Centre National d’Agroécologie (CNA) est un projet porté par des acteurs de l’agroécologie qui y fédèrent leurs compétences et expertises : diffusion et échanges de savoirs, formation et accompagnement technique, outils d’évaluation et de mesure pour concevoir et piloter les systèmes agroécologiques. La raison d’être du CNA est de faire émerger des innovations basées sur les principes de l’agroécologie, au service de l’agriculture et des agriculteurs, pour aider à l’évolution et à la diffusion de leurs pratiques.
Produire durablement une nourriture de qualité
Sécuriser le revenu des agriculteurs
Couvrir et nourrir le sol pour protéger l’environnement
Atténuer le changement climatique
Stopper le gâchis d’énergie et ressources
Produire de la biodiversité
Limiter les pollutions systémiques
Créer du sens et de la solidarité
Copier la nature Toujours couvert/jamais travaillé
Recherche et formation …..sont des bases solides pour l’avenir
Produire et redonner du pouvoir aux agriculteurs ? C’est le pari de Lucien Séguy, « agronome », mais surtout « paysan ». Lucien Séguy dédiera sa vie à la fertilité et au « génie végétale ». Sans dogmatisme, il associera la plus grande diversité biologique possible et les techniques agro-industrielles de pointes, créant lui-même plusieurs variétés de semences de riz à haut-rendements.
Les militants de Solidarité & Progrès ont dénoncé une politique de la famine depuis la fin des accords de Bretton Woods (1971) et la publication du rapport « Halte à la croissance » (1972). Depuis lors, la dérégulation met tous les agriculteurs en compétition, accroissant les inégalités ; et l’idéologie décroissante des institutions financières s’oppose à l’équipement de l’agriculture, même là où elle est la seule ressource.
Est-il encore possible (écologiquement ? ) d’augmenter la productivité agricole ? Dans ce dossier, nous vous proposons de suivre le chercheur Lucien Séguy, à la découverte d’un monde longtemps passé inaperçu, celui des sols vivants. Nous verrons comment l’alliance des sols et des agriculteurs peut vaincre la pauvreté et la faim.
Biographie : Une vie au service de la science ; la science au service des paysans
Lucien Séguy en mission d’appui au Maroc. Source : Fert
Né en 1944 dans une famille paysanne de Dordogne, il est le seul de sa fratrie à poursuivre des études supérieures. Diplômé de l’Ecole Nationale Supérieur d’Agronomie de Toulouse, il se spécialise dans la science des sols (pédologie), puis part en service civique au Sénégal en 1967, développer la riziculture paysanne dans la région de la Casamance. Le Centre de coopération internationale en recherche en agronomique pour le développement (Cirad) lui confie sa première affectation professionnelle au Cameroun. Il y met en évidence l’importance de la gestion du sol pour lutter contre la pyriculariose, un champignon qui détruit les feuilles du riz.
1977 : repéré pour ses travaux sur les cultures de riz, il est appelé au Brésil. Débute sa véritable aventure scientifique. Il y travaillera jusqu’à sa retraite en 2009. En parallèle, il lance des missions d’appui aux paysans dans une dizaine de pays, dont Madagascar, la Côte d’Ivoire, le Gabon, le Cameroun, le Sénégal, la Tunisie, le Viêtnam, le Laos, le Cambodge, le Canada, la France métropolitaine et d’outre-mer, jusque dans les mois qui précèdent son décès en avril 2020.
Le texte qui suit est adapté librement du rapport de Lucien Séguy, son ami Serge Bouzinac, et leurs collègues de la recherche brésilienne : « La symphonie inachevée du Semis Direct dans le Brésil central », rédigé en 2008 pour le Cirad et l’Embrapa (la Société Brésilienne de Recherche Agricole).
Les effets spectaculaires de l’érosion en Europe. Source : agriculture-de-conservation.com / C. Henricot
Lorsque Lucien Séguy arrive au Brésil, il rencontre des paysans ruinés par la dégradation du sol. En cause, le labour.
Le labour est pourtant une méthode qui a fait ses preuves en agriculture. Depuis sa découverte dans le croissant fertile, il y a 10 000 ans, plusieurs civilisations agraires y ont eu recours, de la Mésopotamie à l’Europe et l’Asie. Le labour a permis à des centaines de générations de se nourrir, jusqu’à nous.
Il empêche l’installation des mauvaises herbes et des maladies du sol. Mais exige d’énormes quantités de labeur des paysans. Avec un risque de désertification, particulièrement rapide sous les tropiques.
En climat tropical, les sols ont un aspect minéral, comme la terre de remblai peu fertile de nos terrains vagues. Seuls les cinq premiers centimètres de surface sont noirs et fertiles, comme le terreau ou le compost.
Cette mince couche ne peut s’épaissir, car l’humidité et la température tropicale accélèrent tous les processus du vivant : la croissance des plantes cultivées, comme la décomposition de la matière morte au sol. La « matière organique », noire et fertile, est donc fragile sous ces latitudes.
Dans ce contexte, la technique du labour, qui apporte beaucoup d’oxygène au sol, accélère la décomposition de la matière organique, par les microorganismes du sol. C’est « l’érosion invisible ». De plus, mis à nu et cassé en mottes, le sol est emporté dans les fleuves par les puissantes pluies tropicales. C’est « l’érosion visible ».
En forêt, rien ne se perd !
Tout comme Louis Pasteur, Lucien Séguy a passé une partie de sa jeunesse à peindre, exerçant et développant ses talents d’observateur. En grand scientifique, il interroge sans cesse la nature : « Mon maître, mon juge, mon centre d’inspiration principal, c’est la nature, sous sa complexité la plus grande. Vous avez une question à poser ? Posez-la à la nature. Et elle vous répondra ! Il faut simplement faire quelques manip’ pour qu’elle vous réponde. Et le faire de manière scientifique pour comprendre pourquoi cette réponse ; ce qu’elle veut dire exactement. »
Le but de Séguy est d’inventer une agriculture inspirée de la fertilité naturelle des forêts. Il fonde les principes de cette nouvelle agriculture sur quelques observations.
Lucien Séguy note le rôle essentiel de la « matière morte » dans la fertilité des forêts tropicales. Les « éléments fertilisants » qui s’enfonceraient dans le « sol minéral » pouvant malgré tout être recyclés par les racines profondes.
– Une première observation : Le sol forestier n’a pas besoin d’être labouré pour être meuble. Les organismes du sol forestier et les racines des plantes brassent la matière et produisent la porosité nécessaire à la présence raisonnable d’oxygène et d’eau dans le sol forestier.
1er principe : Lucien Séguy propose donc de semer directement les graines dans le sol, sans labour. L’idée n’est pas nouvelle. Ce semis dit « direct » avait permis aux civilisations précolombiennes de prospérer. La méthode en fut oublié, avec leur effondrement. Cette technique est redécouverte aux Etats-Unis dans les années 60, pour parer aux tempêtes de sable ; suite au « Dust Bowl » des années 30, lors de la grande dépression, qui a profondément meurtri le pays, comme l’illustre le film Les raisins de la colère.
C’est le glyphosate, bêtement diabolisé par des écologistes radicaux, qui a permis aux Etats-Unis d’abandonner le labour, donc d’éviter le désert… Le glyphosate agit en bloquant la photosynthèse des mauvaises herbes, rendant caduc l’archaïque labour. À ce titre, Lucien Séguy propose avec humour que soit décerné « le prix Nobel de la paix à Monsanto, pour sa contribution à la préservation des sols ».
Mais – attention – Lucien Séguy est lucide quant au risque de destruction de l’agriculture par les biotechnologies, quand elles sont développées dans un but mercantile. Au sujet de l’herbicide de Monsanto et des OGM, il nous dit : « la productivité n’a pas augmenté depuis leur adoption massive [alors] que les doses d’herbicide sont plutôt en croissance pour contrôler les [plantes] devenues résistantes au glyphosate. » Avoir recours à de plus grands volumes, inutilement… le contraire de l’écologie !
En Semis Direct, le sol n’est pas retourné : à l’avant du tracteur, un « rolofaca » ou « rouleau hacheur » couche le couvert végétal au sol ; à l’arrière, le semoir sème directement les graines sous cette litière. Source : paysan-breton.fr / Toma Swan
– Lucien Séguy fait une deuxième observation : Les sols forestiers sont couverts en permanence d’une litière végétale. Sous cette litière, les premiers centimètres du sol sont le siège de la fertilité de la forêt tropicale. En profondeur, les éléments nutritifs se font beaucoup plus rares !
2nd principe : Pour avoir un sol aéré, riche en nutriments et protégé contre les pluies diluviennes, Lucien Séguy comprend qu’il doit restituer une partie de la culture au sol. L’enjeu est notamment de nourrir les vers de terre et leur cortège d’êtres vivants. Il choisit de recourir à de « puissantes biomasses » pour couvrir le sol et le régénérer en matière organique, qui confère sa fertilité au sol.
Semis direct dans un couvert de sorgho en Europe. La densité et la vigueur du sorgho en font une excellente « plante géante » de couverture, pour contenir les autres plantes et nourrir le sol. Source : agriculture-de-conservation / Cécile Waligora.
– Une ultime observation permet à cet « ingénieur du végétal » d’identifier la complémentarité des plantes dans les systèmes forestiers : toutes ne prospectent pas l’eau aux mêmes profondeurs ; et certaines plantes peuvent arracher des éléments minéraux au sol, pour les mettre à disposition des plantes de la culture suivante.
3ème principe : Lucien Séguy commence donc un travail qui est au cœur de ce qu’il nomme sa « symphonie inachevée » : Il recherche les meilleures successions et associations végétales, permettant de réduire les dépendances aux fertilisants et aux pesticides, et de maximiser les revenus des agriculteurs.
En rupture avec la monoculture, il propose : « L’incorporation de biomasses de couverture encore plus puissantes et plus diversifiées pour que les couverts végétaux des SCV gagnent en multifonctionnalité gratuite. »
Entre deux cultures commerciales, des couverts végétaux sont semés pour réorienter la biologie, la chimie et la structure du sol. Ici, un mélange de tournesol, phacélie et pois. Source : Ver de terre production
Lucien Séguy appelle sa méthode : « Semis sous Couvert Végétal » (SCV).
Avec son binôme Serge Bouzinac et ses partenaires de la recherche et développement brésiliens, Séguy met en place une méthodologie de « recherche-action » sur « système de cultures pérennes » : le chercheur fait ses essais avec, pour et chez les agriculteurs, sur plusieurs années, pour adapter ses travaux aux contraintes de la production.
Pour ces travaux, Lucien Séguy est nommé docteur « Honoris causa » de l’université de Ponta Grossa, une des meilleures universités agricoles du monde, et “Grand Citoyen” par l’Assemblée Législative de l’Etat du Mato Grosso. Mais le paysan-chercheur tient à rendre hommage aux efforts des agriculteurs brésiliens et des acteurs institutionnels :
« Le Brésil a montré au monde sa capacité à développer en moins de 30 ans une agriculture de conservation que le monde entier admire et lui envie, même si elle est largement encore perfectible comme l’ont démontré nos travaux sur les innovations SCV inspirées du fonctionnement de l’écosystème forestier. »
Ajoutons que, bien que fortement mécanisés, les agricultures du Brésil, de l’Argentine et des Etats-Unis restent peu productives, car peu irriguées : dans la région du Mato Grosso entre 2001 et 2007, les rendements par hectare sont en moyenne : de 3,5 tonnes pour le coton, 2,9 tonnes pour le soja et 3,4 tonnes pour le maïs. En France, le blé produit 7 tonnes par hectare et le maïs 9 tonnes. Un ami et disciple de Lucien Séguy, Christian Abadie, produit même 14 tonnes de maïs dans le sud-ouest de la France !
Pourquoi les agriculteurs des Etats-Unis, de l’Argentine et du Brésil acceptent-ils de produire si peu par hectare ? La réponse est en partie économique.
Regardons plutôt, pour le moment, les résultats de la méthode du « Semis sous Couvert Végétal » de Lucien Séguy.
Bilan des « Semis sous Couvert Végétal »
Amélioration du statut social des paysans, accomplissements économiques et environnementaux divers ; les résultats présentés ci-dessous sont non exhaustifs, mais sont l’occasion d’illustrer les problématiques de l’agriculture.
Résultats du groupe agro-industrielMAEDA : Augmentation de la productivité du soja de 25%, et du coton de 45%. Soit une productivité respectable de près de 4 tonnes de soja par hectare et 5 tonnes de coton. Passage à 3 cultures sur 2 ans au lieu de 2. Les marges ont été multipliées par 3. Le nombre de machines agricoles a été réduit de moitié. Le nombre de prestataires de services a chuté de 71%. Et la consommation de carburant a diminué de 70%. Produire plus avec moins !
Des engrais renouvelables : Le manque d’azote et de phosphore dans le sol est ce qui limite le plus la productivité des champs. L’azote constitue 78% de l’air que nous respirons. Il est obtenu industriellement par la réaction du gaz naturel avec l’air. Quant au phosphore, il est aujourd’hui extrait des carrières de « guano », sites de déjections des oiseaux marins, ressource peu renouvelable.
A titre d’exemple, Lucien Séguy introduit la plante sauvage Stylosanthes, pour sa capacité à fixer l’azote atmosphérique et à mobiliser le phosphore du sol. Cet exploit naturel est dû à des symbioses racinaires, avec des bactéries pour l’azote, et avec des champignons pour le phosphore.
En plus de la Stylosanthes, il choisit la Brachiaria, pour ses racines profondes qui récupèrent les nutriments perdus, de même que pour la densité de son couvert au sol et pour sa forte productivité. Lucien Séguy sème ces deux plantes entre les pieds du maïs qui précède la culture de soja. Il en tire une hausse de productivité pour le maïs et le soja, et s’en sert comme fourrage pour l’élevage.
Une réduction des dégâts : Sur soja, cette fertilité organique permet de réduire le nombre de traitements fongicides des 2/3, voire totalement, et d’obtenir des grains de 1 à 3% plus riches en protéines.
Sur coton, en réduisant de 30 à 50% les apports chimiques d’azote et de potassium, en simple complément de la fertilité naturelle du sol, les dégâts liés au ravageur sont nettement plus faibles. Les passages de pesticides peuvent alors être réduits d’1/3.
Une auto-épuration naturelle : Le sol vivant est un « biodigesteur » des résidus végétaux, mais aussi des pesticides, qu’il réduit en molécules de plus en plus simples. Plus il y a de matière végétale au sol, plus les pesticides restent piégés longtemps dans ce biodigesteur, et donc meilleure est la destruction de ces polluants.
En culture de coton, qui exige souvent de nombreux traitements chimiques, les Semis sous Couvert Végétal (SCV) permettent de réduire le volume d’insecticides de près d’un tiers et le nombre de molécules différentes de 18 à 5. Le volume de fongicides (contre les champignons) est réduit de moitié. La fonction auto-épuratrice des sols vivants permet de réduire sous le seuil de détection l’ensemble des 150 molécules et sous-produits polluants recherchés.
Une valorisation du territoire : Vous voulez sauver la forêt amazonienne ? 1/5ème des surfaces déboisées de l’Amazonie sont à l’abandon, car dégradés par le travail du sol. Les SCV peuvent régénérer ces 16,5 millions d’hectares :
« Les SCV les plus puissants permettent, en 3 à 5 ans, de retrouver les teneurs en matière organique et les caractéristiques de distribution des tailles d’agrégats des sols originels sous forêt. »
Autres sols valorisables : les savanes arbustives du Brésil, appelées Cerrados, réputés incultes. Ces terres, qui rappellent nos garrigues méditerranéennes, offrent plus de 50 millions d’hectares faciles à cultiver en SCV.
Avec la régénération des sols forestiers détruits et la valorisation des sols de savane incultes, Lucien Séguy se réjouit que le Brésil puisse doubler ses surfaces de production sans détruire la forêt amazonienne !
Le bocage Normand. Les haies et les arbres créent un microclimat à l’échelle des parcelles. Auteur : Bournagain.
Enfin, l’eau étant un élément essentiel à la vie, Lucien Séguy recommande de maintenir un maillage continu de végétation native de 20 à 50 mètres autour des parcelles, plutôt que des îlots forestiers. Cette « forme géométrique du défrichement » permet d’éviter l’ « effet de poêle surchauffée qui engendre des courants ascendants d’air chaud qui rejettent les pluies vers les forêts galeries à la périphérie ».
Une symphonie “inachevée”
Comme expliqué précédemment, les agricultures de hautes technologies du Brésil, de l’Argentine et des Etats-Unis pourraient assez facilement produire 1 tonne de plus par hectare. Lucien Séguy déplore le manque de formation technique et pointe aussi des raisons économiques.
À l’image des agriculteurs des Etats-Unis, les agriculteurs du Brésil ont adopté le non-labour, poussés par l’érosion des sols et par les prix très bas payés aux productions agricoles. Le non-labour leur permet de réduire leurs dépenses en matériel et essence, et apporte de la durabilité à leurs exploitations.
Au Brésil Central, où 95% des agriculteurs pratiquent le Semis Direct (1er principe : non-labour), le Semis sous Couvert Végétal (2ème & 3ème principes : une végétation puissante et diversifiée) n’est en réalité pratiqué que sur 30% des champs de soja et 15% des champs de maïs.
Ainsi, depuis les années 2000, étant insuffisamment régénérés, les sols du Brésil Central se compactent et un travail superficiel du sol redevient nécessaire. La méthode alors utilisée est le « discage » des 5 premiers centimètres, ce qui constitue une régression vers le labour et la chimie, des surcoûts et une perte de durabilité.
C’est que produire les couverts à aussi un coût ! Les systèmes de SCV nécessitent des investissements de 11% à 29% supérieurs aux simples systèmes sans labour. Hélas, en plus des aléas naturels, le libéralisme rend les prix payés aux producteurs extrêmement fluctuants et généralement plutôt bas. Ainsi, il est probable que les lendemains incertains découragent l’épargne dans le sol.
La formation professionnelle et des marchés organisés apparaissent comme les deux leviers pour qui veut bâtir une politique du sol vivant.
A la frontière des travaux de Séguy, plusieurs disciplines peuvent être convoquées :
Une myriade de naturalistes (dont les spécialisations sont en proportion de la créativité de la Nature) et d’écologues (spécialisés, eux, dans l’étude des interactions naturels) : mobilisés auprès des agronomes (à recruter également !) pour rechercher de nouvelles plantes de couvertures et de nouvelles stratégies d’assemblage du vivant.
Encore une question de moyen ! L’étude des systèmes naturels nécessite des moyens logistiques que l’approche statistique et la modélisation informatique, généralement privilégiées aujourd’hui en écologie scientifique car moins onéreuses, ne devrait qu’appuyer !
Comme pour les plantes que nous mangeons et qui ont été sélectionnées pour être productives, les sélectionneurs peuvent améliorer les caractéristiques des plantes de couverture : Le sorgho blanc sans tanins à haute teneur en protéines, pour farines, bière, papier et amylose ; le sésame qui contient de 50 à 55% d’huile de qualité supérieure, pour les cosmétiques et l’aviation ; le sarrasin, pour les farines sans gluten, …
L’acclimatation de nouvelles plantes de cultures est aussi un moyen d’améliorer la santé du sol. Par exemple, l’intégration du riz pluvial dans les rotations avec les cultures de soja et coton permettrait de lutter contre les maladies à champignon. Pour rendre cette culture de riz pluvial attractive, le défi est actuellement de mettre au point des variétés à hauts rendements.
Quant aux biologistes et aux généticiens, leurs connaissances à l’échelle cellulaire et moléculaire est une aubaine pour l’étude du sol vivant :
Le séquençage génomique, d’abord, pour l’étude de la diversité microbienne. Champignons et bactéries cachent d’innombrables possibilités de symbioses et d’effets antibiotiques naturels. Il s’agit de tirer parti du haut potentiel des sols vivants !
La recherche dans les OGM aussi, à condition d’être au service de l’agronomie, seule discipline globale des champs : « Les SCV offrent une biodiversité fonctionnelle très efficace […] dans un tel contexte, les OGM pourraient être des auxiliaires précieux, légitimes et incontestablement valorisés pour seulement compléter les services écosystémiques majeurs des SCV. »
Nourrir les BRICS
Lucien Séguy, qui dénonce volontiers le conservatisme français, ne s’étend pas sur la politique du Brésil. Tentons, par nous-même, de comprendre cette très jeune puissance agricole :
L’Embrapa, entreprise d’Etat, a acclimaté le blé, le soja, le maïs et le coton, aux sols et aux climats brésiliens. Son existence depuis 1972 inscrit le Brésil dans une démarche scientifique à long terme et ouvre la possibilité de transferts de compétences avec l’Afrique.
Tracés envisagés du chemin de fer “bi-océanique”.
Bien que le Brésil ait su défendre ses intérêts agricoles devant l’Organisation Mondiale du Commerce, le pays tente dès 2003 de remettre en cause les règles arbitraires imposées par les institutions dévoyées de Bretton Woods. En 2014, au sommet de Fortaleza, est lancée la Nouvelle Banque de Développement des BRICS (Brésil, Russie, Inde, Chine et Afrique du Sud).
Il s’agit pour les pays émergents de réduire leurs coûts de production, non par le dumping, mais par l’apport technologique et l’aménagement du territoire. Cette politique est aussi connue sous le nom de « Nouvelles Routes de la Soie ». Un exemple de coopération agricole entre la Chine et le Brésil : la construction de la voie ferrée bi-océanique, traversant l’Amérique latine d’Est en Ouest. Projet gagnant/gagnant, puisqu’il désenclave l’Amazonie et évite le passage par le canal de Panama pour fournir la Chine en denrées agricoles.
En France, les réseaux de Lucien Séguy ont débuté 30 ans après le bio, mais 15% des surfaces agricoles françaises sont déjà cultivées en Semis-direct, soit le double de l’agriculture biologique. En dehors des radars de la plupart des médias.
Bien sûr, le bio nourrit aussi les gens et doit être soutenu à ce titre. Notons seulement ses limites pour mieux nous détacher de l’agribashing : Une productivité moindre et un sol moins riche en vie.
Apprécions aussi les vrais mérites du bio : une production fraîche, car locale, et des circuits-courts, pour que la valeur aille aux producteurs.
Lucien Séguy entouré de Français passionnés. Source : festival Paysage in Marciac, 2018.
Les réseaux français de Lucien Séguy tentent toutes les dénominations pour se faire connaître du public : agriculture de conservation, agriculture de régénération, agriculture sur sol vivant. Et le ver de terre comme symbole de la méthode.
Ces agriculteurs sont présents dans tous les syndicats. Et la Coordination Rurale, second syndicat agricole de France, entre la FNSEA et la Confédération Paysanne, organise depuis 25 ans des festivals de non-labour.
Un ministre, le socialiste Stéphane Le Foll, a rencontré les leaders de ces mouvements en France, mais son cabinet n’a rien trouvé de mieux à leur proposer qu’une tribune à la COP21. Car la priorité du ministère est d’atteindre la neutralité carbone de notre agriculture, d’ici à 2050. Notre article « Ces milliardaires « écolos » qui veulent nous affamer » dénonce cette politique agricole malthusienne.
Une rencontre fortuite nous avait d’ailleurs permis d’interroger le ministre sur la détresse des agriculteurs et les prix trop bas. Nous lui demandions de mettre en place une limitation des importations et un stockage des surplus pour hausser les prix. Réponse du ministre : « Nous ne sommes pas communistes ! » Ce fut l’occasion pour nous d’évoquer la méthode des trente glorieuses : « Monsieur le ministre, De Gaulle non plus ; pourtant, c’était sa politique ».
Stéphane Le Foll a tout de même soutenu la création de « ver de terre production ». Cette chaîne de diffusion de contenus sur Youtube est animée par des agriculteurs. Mais les médias traditionnels restent en roue libre… et le public avec eux. Le pays a-t-il besoin d’une chaîne de télévision publique, qui soit dirigée par les instituts scientifiques ?
Parmi les mesures pour encourager l’agriculture sur sol vivant, rappelons l’importance d’autoriser le glyphosate pour maîtriser les couverts végétaux.
Enfin, l’hexagone gagnerait beaucoup à mettre en place une taxe sur les importations de soja. Elle inciterait le retour de la production de légumineuses en France, comme durant l’année 1973 (lorsque les Américains ont organisé un embargo du soja contre l’Europe).
Qu’est-ce qu’une légumineuse ? Une légumineuse est une plante qui, comme le pois, la lentille ou le soja, est capable de fixer l’azote présent dans l’air. Elles permettent de fertiliser un champ pour la culture suivante. Les légumineuses servent à diversifier les cultures pour limiter l’installation de maladies du sol. Et leurs graines, riches en protéines, augmentent l’autonomie de l’élevage. Cette taxe est donc une mesure de « protectionnisme éducateur », dans la tradition de l’économiste Friedrich List.
Le Brésil a prouvé qu’il est capable d’entendre cet argument, à condition que son droit au développement soit pris en compte. Retrouvons l’état d’esprit des coopérants, comme Lucien Séguy.
Antoine Beils, mai 2021
Annexe I : De l’importance de la productivité « physique » pour nourrir le monde
L’économiste et homme politique américain Lyndon Larouche a montré la relation entre d’une part, la productivité par travailleur, unité de surface et volume de matières utilisées (les 3 paramètres de la productivité physique) contraints par les handicaps naturels d’un territoire ; et d’autre part, le potentiel démographique de ce même territoire. Dit autrement : Le nombre d’humains que la Terre peut porter n’est pas déterminé par les ressources naturelles, mais par l’efficacité de leur utilisation.
Examinons la productivité physique de l’agriculture.
L’agriculture raisonnée, l’agriculture de précision et l’agroécologie permettent toutes trois de produire autant, avec de moindres volumes de produits chimiques. Ce qui n’est pas forcément le cas du bio, qui a globalement recours à de grands volumes de pesticides « naturels ».
Même constat, avec la faible production du bio par hectare et par travailleur ; bien que l’émergence de pratiques agroécologiques (lorsqu’elles sont compatibles avec le cahier des charges du bio) permettent quelques améliorations.
L’Organisation des Nations Unies (ONU) et sa branche pour l’Alimentation et l’Agriculture (FAO) soutiennent cette productivité physique. Elles parlent d’ailleurs d‘ « agriculture écologiquement intensive » ; mais sans moyens financiers à long terme pour promouvoir les semences à hauts rendements, la mécanisation et les autres moyens logistiques du stockage, du transport et de la chaîne du froid. D’ailleurs, le Programme Alimentaire Mondial des Nations Unies cherche encore 5,5 milliards de dollars, pour l’aide alimentaire d’urgence en 2021.
L’écologie, c’est quand les gros mangent les petits ?
Mais si les prix tombent trop, les agriculteurs renoncent à lutter contre les maladies. Au risque de perdre la production…
C’est aussi le risque pris par le bio, qui, en renonçant à la chimie de synthèse, produit globalement un tiers moins de nourriture que l’agriculture conventionnelle à l’hectare, mais est mieux valorisé commercialement.
La page du productivisme, tant décrié encore, a donc été tournée de force ; bien que les extrémistes verts ne s’en satisfassent pas et exigent l’abandon total des équipements motorisés et de la chimie.
Et le pouvoir de nourrir change de main :
1/ Dépossession des agriculteurs. Puisqu’il faut acquérir du foncier, il faut s’endetter au-delà de ce qui est remboursable. Un problème pour la transmission aux jeunes, mais pas un problème pour le rachat par des fonds financiers.
2/ Concentration de l’agro-industrie. À l’image des abattoirs géants, qui imposent leurs conditions commerciales aux éleveurs.
Annexe II : Puit de carbone & pompe à fric
En climat tropical, le Semis Direct sous puissante biomasse permet de produire de 23 à 32 tonnes de matière sèche, aérienne et racinaire, par hectare. Or, cette matière sèche est faite pour moitié de carbone. Si l’intégralité de cette biomasse est restituée au sol, alors 900 kilos à 2 tonnes de carbone sont séquestrés dans ce sol chaque année, et jusqu’à une dizaine d’années de suite.
Comme développé dans le dossier « Le New Deal vert : sortir du piège de la finance verte », la finance verte tente de mettre les États sous tutelle au nom du climat. Pour l’agriculture, aussi, accepter l’agenda “zéro carbone” est dangereux, car cet agenda mène à l’abandon des énergies fossiles, de la fertilisation azotée et de l’élevage, pourtant nécessaires pour nourrir correctement 9 milliards d’êtres humains en 2050.
Annexe III : Une plongée dans le sol vivant
Voir la vidéo : « Lucien Séguy, semis direct sur 20 millions d’hectares ». École Nationale Supérieure Agronomique de Toulouse (ENSAT).
Arnaud Beils says:Merci pour cet excellent article où j’ai appris beaucoup de choses sur l’intérêt du Semis direct sur Couvert Végétal. Je note notamment la capacité de certaines plantes à aller chercher les minéraux en profondeur. A noter également l’importance méconnus de cette pratique en France (15% de notre production !) Peut-être le semis sous couvert végétal permet-il aussi de réaliser des économies d’eau grâce au fait que le couvert végétal conserve l’humidité contrairement à de la terre nue ?Je souhaiterais te poser quelques questions pour comprendre ce que tu as écrit à fond. 1/ Pourquoi le Brésil n’arrive t’il pas au même rendement que la France ? Tu précises qu’ils sont fortement mécanisés et qu’en 2007, nous (la France) arrivons au double du rendement par hectare. Hypothèse, le sol n’est pas encore assez généré ?2/ L’équivalent du semis direct sous couvert végétal appliqué à de faibles surfaces (- de 1 hectare) est il le maraichage sur sol vivant ? Autrement, quels outils utiliser pour produire en SCV sur une faible surface ?3/ peut-on dire que l’augmentation du CO2 dans l’atmosphère (infime précisons le tout de même) est en parti dût au fait que l’on ne restitue pas la matière organique à la terre ? (d’où un argument supplémentaire en faveur du SCV pour capter le CO2)Encore merci pour cet apport intellectuel. Militons pour que la recherche agronomique ait les moyens d’augmenter notre potentiel démographique. Vive l’Humain explorateur du vivant !Reply
antoinebeils says:Merci à toi l’ami,Pour répondre rapidement : 1/ Comment doubler ou tripler la production : – Le Brésil est un pays aux climats contrastés. Le Mato Grosso par exemple, où a travaillé Séguy, est sec une partie de l’année. Il faut donc investir dans l’irrigation. – Une vraie régénération des sols permet les symbioses racinaires avec la vie du sol, pour valoriser la fertilité, donc une meilleur expression des potentialités de la plante. – Précisions : le soja étant 2 à 3 fois plus riche en protéines que le maïs, les faibles rendements sont compensés.2/ Comment appliquer l’ “agriculture sur sol vivant” sur une surface d’1 hectare : – La chaîne youtube “Ver de Terre Production”, animée par des agriculteurs et des chercheurs, permet de se mettre à jour de “l’état de l’art”, du jardin nourricier aux grandes cultures, en passant par l’élevage, l’arboriculture et le maraichage. 😉3/ La fixation du carbone, un argument pour les sols vivants ? Oui, mais… Le paradigme politique “neutralité carbone en 2050” est dangereux, car il implique la suppression de la fertilisation azoté et le démantèlement de l’élevage, comme l’exigent certains financiers fanatiques. Parlons plutôt de “l’agriculture sur sol vivant” comme d’une méthode pour nourrir le monde, bien nourrir, tout le monde !
Il s’agit d’une glycoprotéine hydrophobe tolérant la chaleur que l’on retrouve à la surface des spores et des mycéliums mycorhiziens. La production de cette substance atteint son maximum chez les mycéliums sénescents. Cette substance à décomposition lente comporterait le tiers du carbone séquestré dans les sols de planète. Sa principale fonction porte sur la stabilisation des agrégats, à la façon d’une colle regroupant l’argile, le limon et le sable fin, avec des effets majeurs sur les propriétés physiques des sols. Ce grumelage assure une meilleure pénétration de l’eau, réduisant les écoulements de surface, de meilleurs échanges gazeux et une rétention de l’eau et des minéraux, notamment le potassium. On peut dire que la glomaline est très étroitement liée à la fertilité des sols. On peut observer la glomaline par immunofluorescence sur les mycéliums, les spores et les agrégats de sol.

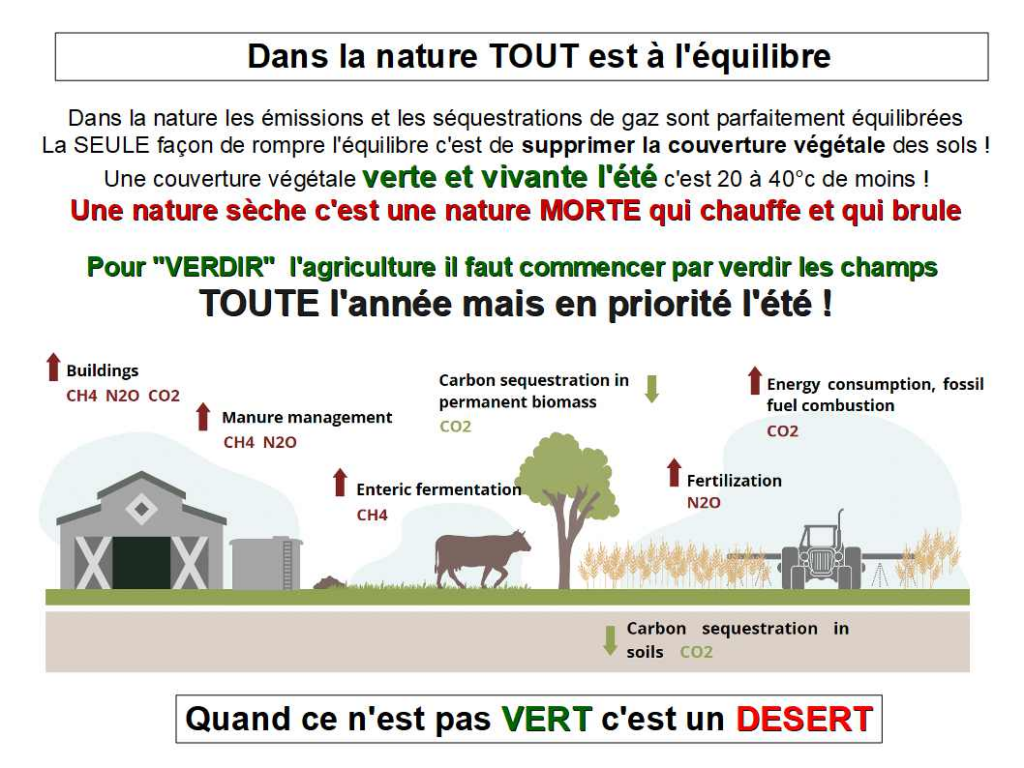

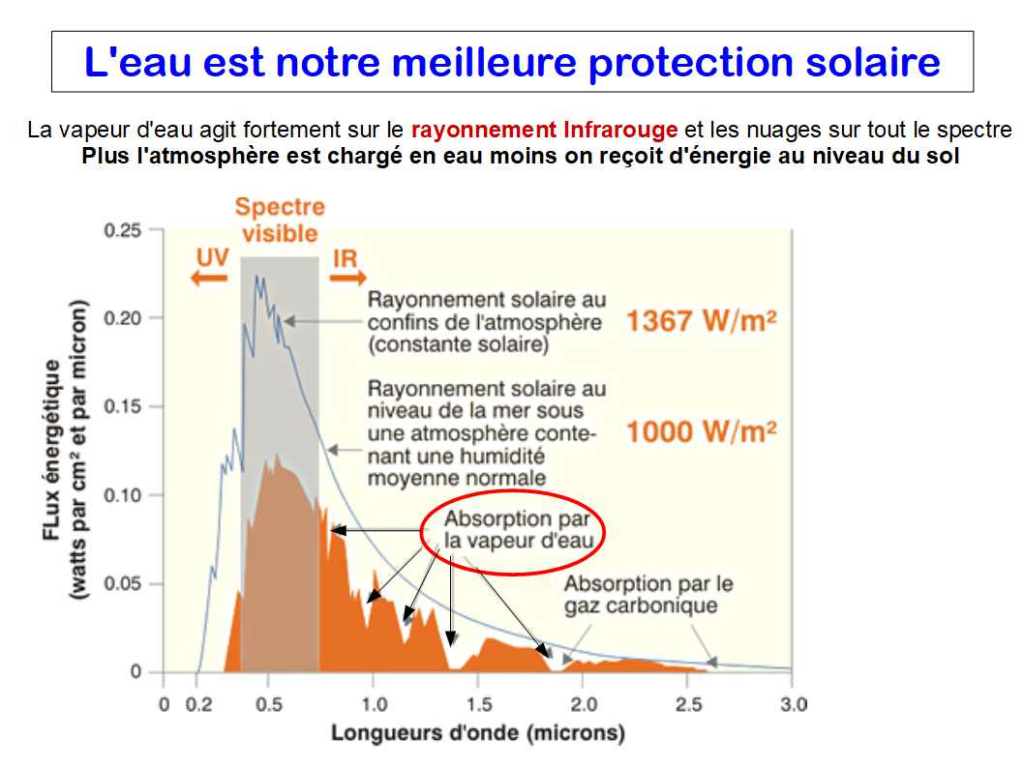



 Ouvrir en mode plein écran
Ouvrir en mode plein écran Ouvrir en mode plein écran
Ouvrir en mode plein écran Ouvrir en mode plein écran
Ouvrir en mode plein écran